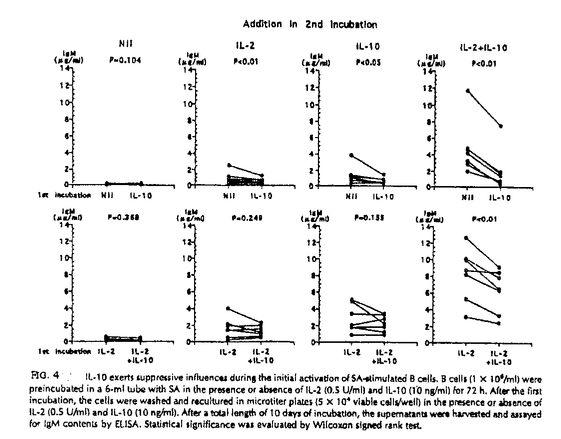
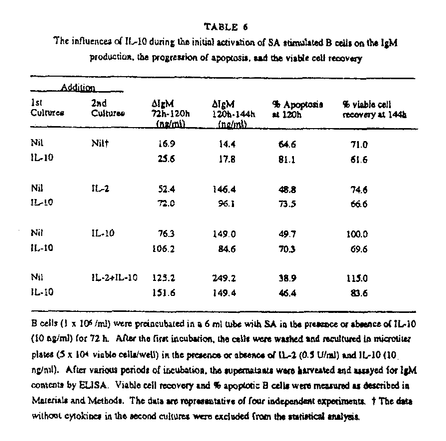
本研究はヒトB細胞の初期活性、増殖、分化の過程において重要な役割を演じていると考えられるIL-10の直接作用とIL-2との共同作用について詳細に検討したものであり、下記の結果を得ている。 1・SA+IL-2刺激B細胞のIg産生に対するIL-10の影響についての検討。 IL-10は単独でも濃度依存性にSA刺激B細胞のIg産生を誘導し得たがその効果はIL-2に比べると微弱なものであった。しかし、IL-2の存在下ではIL-10はSA刺激B細胞のIgM、IgG産生を共に飛躍的に増加させた。この相乗効果は抗IL-2レセプター抗体(抗CD25抗体)、もしくは抗IL-10抗体の添加により阻害された。またIL-10は培養72時間までにB細胞のCD25分子発現を増加させた。さらに、IL-2の作用をほぼ完全に阻害する抗CD25抗体を用いてもIL-10が存在する系ではIg産生を完全に抑えることはできなかった。 2・IL-10がSA刺激B細胞の活性のどの段階でIL-2との相乗効果を現わすのかを検討した。IL-10はSA+IL-2刺激B細胞のIg産生に対し培養開始時や120時間以降に添加するより培養72時間の時点で添加したときに最も強くIg産生誘導作用を示した。一方、IL-2は72時間以降の添加ではわずかなIg産生しか誘導しなかった。IL-2はB細胞の増殖、分化段階ではなく初期活性時に作用すると考えられた。 3・二段階の培養方法を用いてB細胞の活性初期、増殖分化段階におけるIL-2とIL-10の共同作用をさらに詳細に検討した。IL-10はB細胞の初期活性段階の条件にかかわらず72時間以降はIL-2より強いIg産生誘導作用を示した。さらにIL-2とIL-10のB細胞のIg産生に対する相乗的増加作用は活性初期ではなく増殖分化段階で発現した。一方、IL-10が初期活性段階で加えられているとCD25分子の発現増加にもかかわらず後半でのB細胞の反応は抑制された。そしてIL-10の抑制効果はIL-2の添加によって減弱した。 4・IL-10はSA活性化B細胞の生存率を低下させた。PIによるDNA染色では、IL-10の添加によりDNAが断片化した細胞が増加しておりこれはアポトーシスを起こしている細胞の増加を示す。対照的にIL-2の添加ではアポトーシスは減少し、さらにIL-10によるアポトーシスもIL-2により抑制された。これらの結果よりIL-10はSA刺激B細胞のアポトーシスを誘導し、IL-2はIL-10の誘導するアポトーシスから回避させることが示唆される。同時に観察したBcl-2蛋白の発現は、アポトーシス量と逆相関を示した。 経時的な観察では、活性初期よりIL-10を添加したB細胞は72時間以降明確にアポトーシスの増加を示した。 5・72時間までのIL-10の影響がIg産生細胞への分化が終了する120時間での細胞の生存に及ぶかどうかを二段階の培養を用いて確認した。最初にIL-10を添加すると後半加えるサイトカインに関係なく120時間でのアポトーシスが増加し、最初にIL-2を加えるとアポトーシスは減少した。また、IL-10は後半の添加ではIL-2と同程度にアポトーシスを抑制した。さらに詳細に検討すると、培養初期に添加したIL-10は72時間から120時間のIgM産生は増加させたが120時間から144時間のIgM産生を抑制した。さらに120時間から144時間のIgM産生は120時間でのアポトーシスの割合および144時間での細胞の生存率と有為な相関を示した。また、IgGも同様の傾向を示し、この結果がIL-10がクラススイッチを誘導した結果ではないものであると考えられる。 以上、本論文は、IL-10はSA刺激B細胞の初期活性段階ではアポトーシスを誘導するためIg産生を抑制するが、同時にIg産生細胞への分化誘導能を持ち、初期活性以降やIL-2の存在などアポトーシスが抑制された場合Ig産生を増加させることを明らかにし、加えて、IL-2との共存下で観察された複雑な相互作用は実際の液性免疫の発現を理解する上で重要な貢献をなすと考えられ、学位の授与に値するものと考えられる。 | 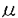 g/mlプロピディウム.イオダイド入りPBSで4℃にて一晩静置し固定、染色。解析にはCoulter Immunology社のEPICS Profile及びEPICS XLフローサイトメーターを用いた。
g/mlプロピディウム.イオダイド入りPBSで4℃にて一晩静置し固定、染色。解析にはCoulter Immunology社のEPICS Profile及びEPICS XLフローサイトメーターを用いた。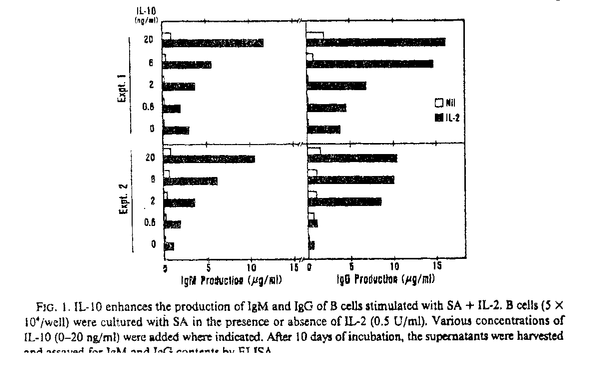
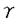 などT細胞が産生するサイトカインや、CD40を介する直接シグナルによりB細胞のアポトーシスが抑制されることが報告され、この様な条件下でIL-10がB細胞のIg産生を誘導するという報告も多い。今回の結果からもIL-10はアポトーシスが抑制された条件下ではIL-2同様、B細胞の分化を誘導できると考えられる。よってIL-2とIL-10のSA刺激B細胞に対する相乗的なIg産生増加作用はIL-10によるIL-2レセプター発現増加と、IL-2によるIL-10のアポトーシス誘導の阻害でIL-2、IL-10それぞれのB細胞の増殖、分化誘導能が発現した結果ではないかと推測される。
などT細胞が産生するサイトカインや、CD40を介する直接シグナルによりB細胞のアポトーシスが抑制されることが報告され、この様な条件下でIL-10がB細胞のIg産生を誘導するという報告も多い。今回の結果からもIL-10はアポトーシスが抑制された条件下ではIL-2同様、B細胞の分化を誘導できると考えられる。よってIL-2とIL-10のSA刺激B細胞に対する相乗的なIg産生増加作用はIL-10によるIL-2レセプター発現増加と、IL-2によるIL-10のアポトーシス誘導の阻害でIL-2、IL-10それぞれのB細胞の増殖、分化誘導能が発現した結果ではないかと推測される。