ニューロトロフィン-3(neurotrophin-3;NT-3)は神経細胞の生存維持ならびに分化促進に必要な神経栄養因子に属する蛋白質である。NT-3は、その一次構造の相同性より、神経成長因子(nerve growth factor;NGF)、脳由来神経栄養因子(brain-derived neurotrophic factor;BDNF)、NT-4、NT-5、NT-6とともにNGFファミリーを形成している。ただし、NT-4とNT-5の構造の違いは種間変異であり、これらは同一の因子と考えられている。これらの因子は、個々の因子が機能するニューロンの種類が異なること、および個々の遺伝子の発現時期ならびに発現組織が異なることから、神経系の構築および構築された神経系の維持にそれぞれが個別の働きを行っているものと考えられている。これらの機能の解析は、従来、ニワトリ胚あるいはマウス、ラット等の哺乳動物のニューロンを初代培養する系で調べられてきたが、これらのin vitroの系では、神経系のような高次生命現象を理解するのに限界があった。近年、遺伝子の生体における機能を解析する手段として、ジーンターゲティングにより遺伝子欠損マウスを作製する手法が開発され、効果を発揮している。この方法を応用すれば、従来不明瞭であったNT-3の生体内の役割が明らかにされると考え、本研究に着手した。 第1章では、ジーンターゲティングを行うためのマウス胚幹(embryonic stem;ES)細胞の樹立について記載した。ES細胞は、その形質が非常に不安定で継代数を重ねたものは劣化しているものが多い。そのため効率よくジーンターゲティングを行うためには独自にES細胞を樹立する必要性があった。ES細胞はマウス初期胚の胚盤胞の内部細胞塊から樹立するが、この方法は技術的に難度が高く、また、多くの胚を同時に扱うのも困難な方法であった。我々は、まず胚盤胞の低い採卵効率を上昇させるため、胚盤胞の1日前のステージである8-細胞期胚を採取することで採卵効率を上昇させ、これを組織培養により胚盤胞へ分化させることにより、一度に多くの胚盤胞を手に入れる系を設定した。次に、胚盤胞の内部細胞塊を得るために、通常は透明帯および栄養外胚葉を、それぞれ酸性タイロード処理ならびに免疫手術で除去するが、これらの操作が必要でないことを確認して省略することにより、作業効率を上昇させた。これら2点の改良点により、従来困難と考えられていたES細胞の樹立を比較的容易に行うことができた。 第2章では、第1章で樹立されたES細胞を用いて、NT-3のジーンターゲティングを行い、NT-3欠損マウスを作製したことについて記載した。NT-3はシグナル配列を含むpre領域、活性を持たないpro領域、および実際に神経栄養因子活性を持つmature領域に分けられる。また、NT-3遺伝子は、そのコード領域が全てひとつのexonに含まれている。そこでジーンターゲティングのためのベクター(ターゲティングベクター)は、このcoding exonのうちmature領域をコードする配列を、相同組み換えにより大腸菌由来のlacZと交換する形のものを作製し、これを用いた。ターゲティングされた場合には、NT-3に代わってlacZがNT-3のプロモーターにより発現されるので、NT-3欠損の解析をすると同時に、NT-3の生体内での発現分布を容易に観察することができた。なおNT-3の発現分布については第4章に詳述した。相同組み換えによりNT-3遺伝子の欠損したES細胞株からキメラマウスを経て得られたヘテロ接合体マウスはNT-3の発現量が半分程度に低下していたものの、正常に生まれ、外見的異常もなく、繁殖能も有していた。このことより、NT-3の発現量が半分に減少することは生体の維持に影響しないことがわかった。ヘテロ接合体マウスを交配して得られたホモ接合体マウスは予測通り、NT-3の発現が完全に消失していた。このNT-3欠損マウスは胎生期に死に至ることなく生まれるが、生まれてきたマウスは四肢及び胴体を強くひねる異常な運動ならびに哺乳不良を呈した。これらのマウスは自然のままで飼育すると生後2日目には、ほとんど全てのマウスが死亡、もしくは母親マウスに食殺された。仮親マウスの下でNT-3欠損マウス4〜5匹に対し正常マウス1匹のグループで飼育すると20%程度のNT-3欠損マウスは離乳させることができた。これらのことから、NT-3欠損マウスの出生直後の致死の原因はNT-3の発現が消失することにより生じたニューロン死が直接引き起こしたものではなく、感覚障害あるいは運動障害が原因の哺乳不良による自然淘汰的なものであると推測された。 第3章では、第2章で作製したNT-3欠損マウスの組織レベルでの異常の解析について記載した。NT-3欠損マウスの外見ならびに行動学的観察より、NT-3欠損マウスは三叉神経系ならびに固有知覚系に障害のあることが推測された。一方、運動神経系においては、マウスが四肢を動かすことが可能なことから重篤な異常は考えられないが、従来の知見より異常が生じている可能性も考えられた。さらに、NT-3欠損マウスの行動がヒトの小脳失調に類似していたこと、ならびにマウス小脳でNT-3の強い発現が認められることから、小脳の変化についても興味が持たれた。以上のことから、NT-3欠損マウスの知覚神経節(三叉神経節、後根神経節)、脊髄および小脳について組織学的解析を行った。その結果、NT-3欠損マウスの三叉神経節では対照マウスと比較して約70%の体積減少が認められた。また、後根神経節においても50-60%の体積減少が観察された。いずれの場合も残存しているニューロンには形態異常は認められなかった。さらに、これらの末梢知覚神経節に対してNGFファミリーの高親和性受容体であるtrkファミリーチロシンキナーゼの発現解析をin situ hybridizationで行ったところ、NGFの受容体であるtrkAと、BDNFならびにNT-4の受容体であるtrkBの発現細胞には変化が認められなかったが、NT-3の受容体であるtrkC発現細胞のみがほとんど消失していることが判明した。このことから、末梢知覚神経節では個々のニューロンがtrkA、trkB、trkCのいずれかひとつを発現していて、それぞれに対応するニューロトロフィンに完全に依存していることが明らかとなった。一方、脊髄においては、脊髄前角の運動ニューロンの形態学的異常ならびに有為な細胞数減少のいずれも認められなかった。また、trkC発現量にも変化は認められなかった。脊髄には、trkファミリーのうちtrkBとtrkCが発現していることが知られているが、この結果より脊髄運動ニューロンはこの両方を発現していて、NT-3欠損マウスではBDNFまたはNT-4によりNT-3の欠損が補われたものと推測された。小脳については、trkCの発現が認められるプルキンエ細胞層ならびにNT-3の発現が認められる顆粒細胞層について形態学的観察を行ったが、NT-3欠損マウスでは、いずれの細胞でも細胞密度、細胞の形態に変化は認められなかった。また、顆粒細胞層の層構造の成熟度にも異常は認められなかった。以上のことを総合すると、NT-3欠損マウスに見られる運動障害は、脊髄、小脳を含む中枢神経系の異常によるものではなく、末梢知覚神経系のうち、固有知覚が欠落したため、マウスが自己の姿勢および歩行時の四肢の位置関係を認識できないために生じたものと推測された。 第4章では、NT-3欠損のためにNT-3遺伝子と相同組み換えで交換した大腸菌由来のlacZを利用して、NT-3の発現分布を観察した結果について記載した。NT-3と置換されたlacZの発現により生じた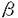 -ガラクトシダーゼ活性をX-Galで染色することによりin situ hybridizationのような煩雑な手順を経ずとも容易にNT-3の発現分布を観察する事ができた。NT-3の発現は、マウス10日胚では、中脳、体節ならびに下顎に観察された。胎生中後期(13日から17日胚)になると中枢神経系での発現は相対的に減少し、末梢神経の標的組織、すなわちwhisker-pad、眼、耳たぶ、内耳、外性器、骨格筋、腸管壁、膀胱壁ならびに動脈壁に強い発現が認められた。これらの発現分布はヘテロ接合体とホモ接合体の間で差は認められなかった。神経系組織では、三叉神経節、後根神経節等のニューロンに発現が認められた。これらの部位でのNT-3発現細胞はヘテロ接合体の方がホモ接合体よりも多かった。従って、ホモ接合体で消失しているニューロンはtrkC陽性ニューロンであると同時にNT-3陽性ニューロンであることが強く示唆された。この結果は、NT-3をはじめとするニューロトロフィンが標的由来型の作用のみならずautocrine型の作用でも働いていると考えられている仮説を支持した。小脳は中枢神経系の中で最もNT-3発現量の多い部位のひとつであるが、小脳内での発現分布については従来報告がされていなかった。NT-3の小脳の部位別の発現分布を検討したところ、小脳の後葉部のlobule VIIからXに局在していることが明らかとなった。また、小脳での発現は3週齢までは上昇するが、その後加齢とともに減少し8ヶ月齢にはほとんど消失した。NT-3欠損マウスのこの部位に組織学的な異常は見いだされなかったが小脳後葉部に特異的なNT-3の発現は興味深い知見である。 -ガラクトシダーゼ活性をX-Galで染色することによりin situ hybridizationのような煩雑な手順を経ずとも容易にNT-3の発現分布を観察する事ができた。NT-3の発現は、マウス10日胚では、中脳、体節ならびに下顎に観察された。胎生中後期(13日から17日胚)になると中枢神経系での発現は相対的に減少し、末梢神経の標的組織、すなわちwhisker-pad、眼、耳たぶ、内耳、外性器、骨格筋、腸管壁、膀胱壁ならびに動脈壁に強い発現が認められた。これらの発現分布はヘテロ接合体とホモ接合体の間で差は認められなかった。神経系組織では、三叉神経節、後根神経節等のニューロンに発現が認められた。これらの部位でのNT-3発現細胞はヘテロ接合体の方がホモ接合体よりも多かった。従って、ホモ接合体で消失しているニューロンはtrkC陽性ニューロンであると同時にNT-3陽性ニューロンであることが強く示唆された。この結果は、NT-3をはじめとするニューロトロフィンが標的由来型の作用のみならずautocrine型の作用でも働いていると考えられている仮説を支持した。小脳は中枢神経系の中で最もNT-3発現量の多い部位のひとつであるが、小脳内での発現分布については従来報告がされていなかった。NT-3の小脳の部位別の発現分布を検討したところ、小脳の後葉部のlobule VIIからXに局在していることが明らかとなった。また、小脳での発現は3週齢までは上昇するが、その後加齢とともに減少し8ヶ月齢にはほとんど消失した。NT-3欠損マウスのこの部位に組織学的な異常は見いだされなかったが小脳後葉部に特異的なNT-3の発現は興味深い知見である。 総括では、NT-3欠損マウスと他のNGFファミリー欠損マウスおよびtrkファミリー欠損マウスとの比較により明らかとなった各因子の特異的な作用について記載した。知覚神経系では各因子の役割は明確に分けることができ、NGFは温冷痛覚、BDNFは触圧覚、NT-3は固有知覚を維持していることが明らかとなった。しかしながら、中枢神経系ではいずれの欠損マウスも明確な異常が認められなかった。ジーンターゲティングの方法論的な改良が試みられており、コンディショナルな遺伝子欠損に成功すればNT-3を含むNGFファミリーの機能がさらに明確になるものと期待される。 |