大脳皮質は、階層的に異なる領野が互いに順行性と逆行性の神経投射によって双方向性に連絡するよう構成されている。しかしながら、これまで単一ニューロンのレベルでは、逆行性結合の機能的役割に関する知見は極めて乏しい。神経心理学的研究から、事実や出来事に関する長期記憶すなわち陳述記憶の形成において、辺縁皮質から新皮質への逆行性結合が重要な役割を果たすとの仮説が提唱されてきた。この仮説は、脳損傷患者に対連合を組んだ一連の言葉や図形を学習させ、その学習能力を検討する方法に基づいている。 対連合学習によって図形に関する連合的な意味のコード化を獲得するニューロン群が、サルの下部側頭葉前腹側部で見出されている。この領野を霊長類の視覚記憶の貯蔵庫とする仮説も提唱されている。そこで我々は、長期記憶の形成に大脳皮質の逆行性入力が、どのような機能的役割を担っているかを検討することにした。 この実験では、単一神経細胞活動記録法と外科的処置を施した慢性のサル標本とを組み合わせた。2頭の日本サルを用いた。サルの片側の嗅皮質(嗅内皮質と傍嗅皮質)のニューロンを神経毒であるイボテン酸で破壊し、辺縁皮質から下部側頭葉ニューロンへの逆向性信号を遮断して、下部側頭葉ニューロンの反応性を調べることにした。この逆同性入力の遮断効果を見るためには、破壊側と非破壊側の下部側頭皮質間の結合を断つ必要がある。両半球の前部側頭葉皮質は、脳梁ではなく前交連を介して互いに連絡している。そこでまず、サルの前交連を切断した。 回復を待って、サルにコンピューターで作った12組24枚の人工図形(セットA)を用いて視覚性対連合記憶課題を学習させた。学習し終ってから、図形に強く選択的に反応するニューロンが高密度に分布する領野を同定して、破壊前の対照記録を行なった。その後、片側の嗅内皮質と傍嗅皮質に、ハミルトン・シリンジ針を用いて格子状にイボテン酸を注入した。一方のサル(M)では22トラックの注入を要し、他方のサル(N)では19トラックを要した。これらの皮質は、同側の下部側頭葉皮質に大量の逆向性投射を送っている。イボテン酸は細胞体を選択的に破壊して、白質を通る神経繊維は障害しないことが報告されている。組織学的検索によって、破壊は嗅脳溝を囲む内側、外側の皮質全体に及び、さらに嗅内皮質と傍嗅皮質のほとんどにも及んでいることが確認された。注入後もサルは手掛かり刺激を正常に固視できることが、磁気サーチコイルによる眼位モニターから確認された。さらに破壊前に学習した対連合図形(セットA)を再学習するとともに、新しい対連合図形(セットB)も学習できた。その後、単一神経細胞活動の記録を破壊前の対照記録と同じ領域から行なった。 手掛かり刺激呈示期のはじめの400ミリ秒間と、これに先立つ固視パターン提示の直前400ミリ秒間の活動電位の数を測定した。両者が統計的に有意に異なれば(paired-t test,P<0.05)、そのニューロンは手掛かり刺激に対して反応性があるとした。また、ある刺激に対する反応がその他の刺激に対する反応よりも有意に強ければ(analysis of variance,P<0.05)、そのニューロンは手掛かり刺激に対して選択的であるとした。破壊前に2頭のサルからサンプルした256個のニューロンのうち、109個は反応を示さなかった。また弱い反応は、最適刺激に対する反応とは考えられないので、最適刺激に対する反応が毎秒20スパイクを越えた92個のニューロンを解析の対象とした。この92個のうち10個は刺激選択性がなかった。破壊後は、363個のニューロンの反応を記録した。このうちセットA刺激を用いて反応性を調べのは341個で、セットB刺激については332個であった。341個のニューロンのうち202個は反応せず、67個は弱い反応しか示さなかった。一方、332個のうち195個は反応を示さず、62個は弱く反応しただけであった。結局、セットAについては72個のニューロンの反応を、セットBについては75個のニューロンの反応を解析した。72個のうち7個、75個のうち8個は刺激選択性を持たなかった。 サンプルした各々のニューロンについて、その対連合をコードする性質を解析するために二つの指標を用いた。一つは標準的な相関係数(CC)で、もう一つはペアー・インデクス(PI)である。相関係数(CC)は、 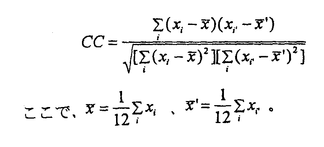 Xiは、i番の図形が手掛かり刺激として提示されたときの平均発火頻度である。i番とi’番の図形はペアーを成している。分母から分かるように、-1<CC<+1である。もう一つの指標は、ペアー・インデクス(PI)で(Sakai,K.& Miyashita,Y.(1991)Nature354,152-155.)、 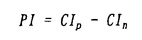 ペアーを組んだ図形どうしについて、 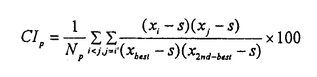 ペアーを組んでいない図形の任意の組合せについて、 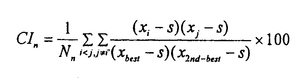 Xiは、i番の図形が手掛かり刺激として提示されたときの平均発火頻度である。i番とi’番の図形はペアーを成している。Sは、自発発火頻度である。Xbestは各々のニューロンがもっとも強く反応した図形に対する平均発火頻度で、X2nd-bestは各々のニューロンが2番目強く反応した図形に対する平均発火頻度である。NpとNnはそれぞれ、2つのインデクスでの図形の組合せの総数を表す。さらに、同じ刺激図形に対する試行毎のニューロン反応の揺らぎを検討するために、反応変動インデクス(response variability index:VAR)を用いた。その定義は、各々のニューロンの最適刺激に対する発火頻度の標準偏差を最適刺激に対する発火頻度の平均値で割ったものである。VARの値が大きいほど、最適刺激に対する試行毎の反応の揺らぎが大きいことを表す。 2頭のサルから得られた結果は、2次元分散分析によってサルの間で統計的な有意差の無いことが確かめられたので、一つにまとめた。嗅内皮質と傍嗅皮質の破壊後、ニューロンの最適刺激に対する発火頻度は、35±2スパイク/秒(セットAで;mean±s.e.m.)と38±2(セットBで)であり、破壊前(31±1)より強かった。しかし、CCは、破壊前(0.166±0.038;mean±s.e.m.)よりも破壊後(セットAで0.024±0.041、セットBで-0.004±0.033)の方が有意に減少した(P<0.05;t-test)。PIも破壊前(1.00±0.33;mean±s.e.m.)に比べて破壊後(セットAで0.04±0.35、セットBで-0.19±0.27)有意に減少した(P<0.05)。自発発火頻度(spontaneous discharge rate:SPON)は、3つのニューロン・グループ間で有意差は無かった(P>0.2)。VARも、3つのグループ間で有意差は見られなかった(P>0.2)。 これらの結果から我々は、霊長類の視覚記憶システムのうち、新皮質の連合野における連合記憶のニューロン・コードの生成は辺縁系に依存し、いっぽう個々の図形パターンのニューロン・コードの生成は辺縁系に依存しないと結論する。 |