食感は、食品の商品価値を決定する重要な因子であるが、無脊椎動物の可食部を対象としたこの方面の研究は極めて乏しい。一般に、生食した場合の魚介類の硬さは、結合組織中に含まれるコラーゲン量に比例することが知られている。ナマコ類体壁は結合組織そのものであり、その生食したときのコリコリした歯ごたえにもコラーゲンが関与することは想像に難くない。一方、生理学的な観点からの研究によると、ナマコ類体壁の伸縮性には、中性塩の影響が大きいことが明らかにされている。この事実は、ナマコ類体壁の強度の発現にはコラーゲンの他に、結合組織を構成する他の細胞外マトリックス成分も関与することを強く示唆するが、その機構は不明のままとされてきた。 本研究はこのような背景の下、マナマコStichopus japonicusを対象に、体壁の強度および微細構造に及ぼす中性塩の影響を調べた。その結果、体壁強度にはグリコサミノグリカン(GAG)が関係することが強く示唆された。そこで体壁から実際にGAGを単離して、その粘度や分子形態に及ぼす中性塩の影響を調べた。さらに当該GAGにつき、酵素消化による分析やメチル化分析を骨子とする方法により、詳細な構造解析を行った。得られた研究結果の大要は以下の通りである。 1.マナマコ体壁の強度および微細構造 マナマコ体壁のホモジネートの貯蔵中、ペプチド態Nが増大しなかったことから、その主要タンパク質であるコラーゲンは貯蔵中に分解せず、したがってマナマコ体壁の死後の急激な軟化がコラーゲンのプロテアーゼ分解によらないことをまず確認した。既に、Eylers(1982)はナマコ類の体壁の伸縮が1価や2価の陽イオンにより著しく影響されることを明らかにしている。そこで本研究では、脱塩処理したマナマコ体壁に種々の陽イオンからなる中性塩を加えた後、直径1mmの円筒状プランジャー付きのサン科学社製R-UDJ-DM型レオメーターを用いて、2cm/minの速度で貫入強度を測定した。その結果、体壁の強度は0.1MのNa+やK+の添加により、当初の値(0.6〜1.4kg)の33〜55%に急減し、さらにNa+では0.3M、K+では0.4Mで強度が見かけ上消失した。一方、Ca2+、Mg2+といった2価の陽イオンも0.1Mまでは軟化作用を示したが、それ以上の濃度でも生時の50%以上の強度が見られ、1価陽イオンの場合と様相を異にした。 1価や2価の陽イオンの影響からマナマコ体壁の強度の発現にはGAGの関与が想定された。まず、通常の透過型電顕観察を行ったところ、マナマコ体壁は若干のコラーゲン線維が無秩序に存在する極めて疎な組織であることが示された。次に、GAGに特異的なルテニウムレッド染色を体壁に施して電顕観察したことろ、コラーゲン線維束以外の空間に、線維状あるいは部分的に網目状のGAG様集合体が存在することが明らかとなった。さらに0.4MNa+処理により軟化した体壁組織をルテニウムレッド法で電顕観察したところ、GAG様集合体がコラーゲン線維束の周囲に凝集することが明らかとなり、GAG様物質の存在がマナマコ体壁の強度を左右することが組織レベルでも示唆された。 2.マナマコ体壁グリコサミノグリカンの単離と物理化学的性状 前述のようにマナマコ体壁の強度や微細構造に及ぼす陽イオンの影響から、その変化にGAGの関与が示唆されたので、これをプロナーゼ処理や四級アンモニウムイオン(QN+)をもつ塩化セチルピリジニウムによる共沈などを骨子とする方法で単離した。170gの体壁から約1gのQN+処理前の粗GAGが得られた。セルロースアセテート膜電気泳動では粗GAGは単一のバンドを示し、体壁中のGAGは一種類であると考えられた。また、同電気泳動におけるQN+処理後の精製GAGの易動度は、各種の泳動溶媒のいずれにおいてもヘパリン(Hep)標品の易動度とほぼ一致したのに対し、コンドロイチン硫酸AやCなど他のGAG標品の易動度とは完全に異なった。したがって、精製GAGはHepと同様に高度に硫酸化されていることが示唆された。カルバゾール法によるウロン酸の定量値に基づき、マナマコGAGの体壁湿重量中の含量は0.03%と算定された。 次に、マナマコ体壁中と同じ濃度の0.03%の精製GAG水溶液を用いて、その比粘度(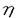 sp)に及ぼす各種陽イオンから成る中性塩の影響を調べた。その結果、当初の比粘度0.70が、各0.1MのNa+、K+、Ca2+およびMg2+存在下でそれぞれ0.47、0.51、0.49および0.57へと減少したが、イオン濃度を0.2M以上とすると、1価陽イオンで引き続き粘度が低下したのに対し、2価陽イオンでは逆に上昇傾向に転じた。これら粘度変化のパターンは、体壁の陽イオン添加による強度変化と高い相関を示した。 sp)に及ぼす各種陽イオンから成る中性塩の影響を調べた。その結果、当初の比粘度0.70が、各0.1MのNa+、K+、Ca2+およびMg2+存在下でそれぞれ0.47、0.51、0.49および0.57へと減少したが、イオン濃度を0.2M以上とすると、1価陽イオンで引き続き粘度が低下したのに対し、2価陽イオンでは逆に上昇傾向に転じた。これら粘度変化のパターンは、体壁の陽イオン添加による強度変化と高い相関を示した。 さらに、1%濃度の精製GAG水溶液につき超遠心分析によって沈降係数(S)を測定したところ、当初1.6Sであったものが、各0.1MのNa+、Ca2+およびMg2+存在下でそれぞれ2.7S、3.6Sおよび3.2Sへと増大した。さらに、精製GAGの蒸留水中の分子量は18,000と計算され、0.1MのNa+存在下で2分子、0.1MのCa2+あるいはMg2+存在下で3〜4分子が会合して大きな分子となることが示された。これらの結果から、GAGがマナマコ体壁の強度発現の原因物質であることが確認された。 3.マナマコ体壁グリコサミノグリカンの構造 マナマコ体壁GAGの構造につき詳細な検討を加えた。まず、前述の精製GAGの化学組成分析を行ったところ、硫酸基、N-アセチルガラクトサミン(GalNAc)、グルクロン酸(GlcA)およびフコースがGAG1g当り、それぞれ3.12、0.78、0.98および1.64mmol存在することが判明した。クロマトグラフィーを取り入れた改良法で精製したGAGもほぼ同様の化学組成を示した。精製GAGのIRスペクトル解析では、比較として用いたスルメイカ軟骨由来のコンドロイチン硫酸E(ChS-E)と全体的によく似たスペクトルを示したが、900および960cm-1の吸収が存在した点で様相を異にした。また、コンドロイチナーゼABCおよびACのいずれによっても酵素消化されず、特異な構造を有することが示唆された。 次いで、精製GAGを0.05M硫酸を用いて80℃で加水分解したところ、GAGは経時的に低分子化するとともに、2種のフコース含有低分子を生成した。精製GAGはコンドロイチナーゼ類で消化されなかったのに対して、部分水解処理を施したGAGは、コンドロイチナーゼACによりほぼ完全に消化され、GalNAcおよびGlcAからなる4種の不飽和二糖( Di-S)を生成した。生成したDi-Sには非硫酸化物のDi-0Sのほか、GalNAc残基のO-4あるいはO-6位が硫酸化されたDi-4SおよびDi-6S、さらにはO-4およびO-6位がいずれも硫酸化されたDi-4,6Sが認められた。4種Di-Sのモル含有率は上記の順に11.2、10.4、56.0、および22.4%であった。なお、Di-4,6Sを含むGAGは、一般的にChS-Eと呼ばれる。以上の結果より、マナマコ体壁GAGは、ChS-Eをコア多糖とし、そこからフコースが枝分かれして伸展するという、極めて特異な構造を持つことが明らかとなった。 Di-S)を生成した。生成したDi-Sには非硫酸化物のDi-0Sのほか、GalNAc残基のO-4あるいはO-6位が硫酸化されたDi-4SおよびDi-6S、さらにはO-4およびO-6位がいずれも硫酸化されたDi-4,6Sが認められた。4種Di-Sのモル含有率は上記の順に11.2、10.4、56.0、および22.4%であった。なお、Di-4,6Sを含むGAGは、一般的にChS-Eと呼ばれる。以上の結果より、マナマコ体壁GAGは、ChS-Eをコア多糖とし、そこからフコースが枝分かれして伸展するという、極めて特異な構造を持つことが明らかとなった。 さらに、フコースの枝分かれ構造の詳細を知るために、精製GAGを化学的に脱硫酸化、カルボキシル基還元した。一方、ヒドラジン分解/亜硝酸分解処理により低分子化GAGをも調製した。こうして得た、精製標品、その脱硫酸化物、脱硫酸化/カルボキシル基還元標品、さらには低分子化した画分の計4標品を、メチル化分析に供した。その結果、マナマコ体壁GAGの枝分かれ構造は、2残基のフコースが 1→3グリコシド結合したもので、フコース分岐とコア多糖の二糖単位はほぼ等モルずつ存在することが判明した。さらに、フコース分岐の約20%はコア多糖のGlcAのO-3位に結合していることが確認された。残りの約80%は、GalNAcのO-4あるいはO-6位に結合していることも強く示唆された。また、各フコース分岐には、硫酸基が平均2.5個結合し、その結合位置の違いによって6種類の分岐構造が存在することも明らかとなった。 1→3グリコシド結合したもので、フコース分岐とコア多糖の二糖単位はほぼ等モルずつ存在することが判明した。さらに、フコース分岐の約20%はコア多糖のGlcAのO-3位に結合していることが確認された。残りの約80%は、GalNAcのO-4あるいはO-6位に結合していることも強く示唆された。また、各フコース分岐には、硫酸基が平均2.5個結合し、その結合位置の違いによって6種類の分岐構造が存在することも明らかとなった。 以上、本研究により、まずマナマコ体壁の強度および微細構造は、中性塩によって著しい影響を受けることが明らかにされた。そこで実際に体壁からGAGを単離し、その粘度や分子形態に及ぼす中性塩の影響を調べたところ、体壁の強度変化の場合と類似の傾向を示した。したがって、マナマコ体壁を生食したときのコリコリとした歯ごたえは、体壁中に約0.03%含まれるGAGの性状に由来するものと結論した。さらにこのGAGの構造を詳細に調べたところ、硫酸化フコース分岐を有するChS-Eといった極めて特異な構造を持つことが示された。これらの成果は、マナマコ体壁の品質向上に有用であり、食品化学に貢献するところが大きいだけでなく、新奇GAGの構造決定において、比較生化学にも資するものと思われる。 |