本論文は、天然セルロースの結晶格子定数がセルロースの起源によって変動する点に注目し、その理由を実験的に追求したもので、天然セルロース結晶はセルロースI (3斜晶)とI (3斜晶)とI (単斜晶)の2つの結晶相がある範囲の比率で複合しているとする、著者の言う2相モデルで統一的に説明できることを示した研究である。論文は8章から成り立っている。 (単斜晶)の2つの結晶相がある範囲の比率で複合しているとする、著者の言う2相モデルで統一的に説明できることを示した研究である。論文は8章から成り立っている。 第2章では海草、バクテリア、木材、サボテン、ホヤの5グループ12種類の精製セルロースの面間隔を測定し、さらに、IをIに転移させる熱処理を行った後、再び面間隔を測定して、セルロースはIの相を多く含む海.藻・バクテリア型とIがほとんどのコットン・ラミー型に分別され、熱処理によって前者はコットン・ラミー型に変態することを示した。 第3章では、系統が進んでいるグループほど、単斜晶角が直角に近いとされる木材セルロースについて検討した。前章で得た海藻・バクテリア型とコットン・ラミー型に分類する判別式に、木材板目板の面間隔を代入したところ、およそ1/3の木材が海藻・バクテリア型と判別された。ところがホロセルロースにした場合は、すべてがコットン・ラミー型に判別された。これより木材セルロースは、I(単斜晶)がほとんどであるコットン・ラミー型であろうと結論づけた。また、木材中に存在する準結晶や非晶成分が、X線の回析ピークをシフトさせるのであろうと推察した。 第4章では、前章で推察した木材中の準結晶や非晶成分の影響を検討するために、ドロノキあて材と正常材のセルロースについて、固体13C NMR、顕微分光FT-IR、制限視野電子回析を加えて検討した。あて材の固体13C NMRスペクトルは、C1とC4の位置に鋭いダブレットの共鳴線が存在し、純粋なIのスペクトルと一致した。一方、正常材のスペクトルには、ダブレットのC1の間にI類似の共鳴線が観察されたが、これは非晶成分の磁化の減衰と一致していたので、非晶由来のピークであると判断した。FT-IRスペクトルには、Iに帰属される吸収バンドが存在しているが、Iに特徴的な吸収バンドは観測できなかった。未木化のG層と木化したS2壁から得た制限視野電子回析ならびにあて材と正常材のX線回析による繊維図には、分解能に差はあるものの、三斜晶に由来する回析点は観測できなかった。すなわち、いずれの手法もIの存在を確証させるものがなかったので、ドロノキのセルロースは細胞壁の木化、未木化を問わずI(単斜晶)であると結論した。 第5章では、セルロースの2相性と植物の系統について検討した。すなわち、海藻から木材に至るさまざまな起源のセルロースを対象に、2相モデルに基づく分類を行い、系統の進んだグループはコットン・ラミー型に、遅れたグループは海藻・バクテリア型に見いだされ、さらに、セルロース合成にかかわるターミナルコンプレックスが直線型のグループは海藻・バクテリア型の、ロゼット型であるグループはコットン・ラミー型のセルロースを産出することを明らかにした。これより、セルロース結晶の2つのタイプは植物の系統と密接な関連があり、両者の境界は緑藻類に存在することを示した。 第6章では、放射光扮末回析ならびに中性子扮末回析法を用いてX線回折強度曲線の分解法を検討した。Gauss関数など6つの関数について信頼度因子を比較し、pseudo Voigt関数が最も良く適合することを示した。 第7章では、X線回折からIとIの混合比を簡便に算出する方法を検討した。その結果、面間隔が0.62nm、0.59nmと0.54nm、0.52nmの二つの複合回析を用いて判別分析を行い、すべての試料がIを多く含むタイプとIを多く含むタイプに間違いなく分類されることと、さらに、高温水蒸気処理シオグサセルロースを用いて、この方法が混合比を推定するのに有効であることを示した。 第8章では、1本のミクロフィブリル中でIとIの相がどのように存在しているかを段階的な加水分解試料の電子回折、電顕観察によって検討した。その結果、シオグサセルロースは外側にIの相を持つ平行構造であると推定した。 以上本論文は、天然セルロースの結晶構造について基礎的な知見を数多く与えたもので、学術上、応用上貢献するところが大である。よって審査員一同は、本論文が博士(農学)の学位論文として価値あるものと認めた。 | 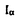 と
と の二つがあり、天然セルロースは
の二つがあり、天然セルロースは (monoclinic angle)=97.08゜であり、既往の2本鎖の単位胞の報告と一致した。以上、いずれの手法からも
(monoclinic angle)=97.08゜であり、既往の2本鎖の単位胞の報告と一致した。以上、いずれの手法からも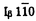 、
、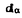 (100)=0.613nm,
(100)=0.613nm, =0.603nm,
=0.603nm, (110)=0.535nm,
(110)=0.535nm,