腎臓の近位尿細管は濾過されたNaClの大半を再吸収しており、この過程は体液恒常性の維持などに重要な役割をはたしている.このうちNaイオンの細胞内輸送については解析が進み、その調節機構も明らかにされつつある.ところが、Clイオンの細胞内輸送機序については不明の点が多く残っている.特にこの部位の基底側膜については今までの数多くの電気生理学的な解析により有意なClコンダクタンスは存在しないとされてきた.従って、基底側膜を通過するClの輸送は電気的に中性な輸送経路によると考えられてきており、とりわけK-Cl共輸送体の存在が提唱されてきた.しかしこの報告は管腔側からの通電という疑問の残る手法を用いたものであり、また小胞を用いた実験からは基底側膜にClコンダクタンスが存在する可能性も示唆されている.さらに最近の分子生物学的な解析によれば,近位尿細管には幾つかの異なるClチャンネルのmRNAが分布していることが明らかになってきた.我々は今回これらの点を踏まえ、比較的単純な輸送特性を持つ近位尿細管終末部(S3部)において電気生理学手法を用い、基底側膜におけるClの輸送機序について再解析することにした. 管腔をcollapseさせたウサギ単離近位尿細管S3部において溶液のk濃度を増加させると、図1に示すように細胞内電位(Vb)が脱分極するとともに細胞内Cl濃度([Cl-]i)は漸増し、またこの反応はCl/HCO3交換輸送体を完全に阻害する濃度のDIDSにより抑制されなかった. 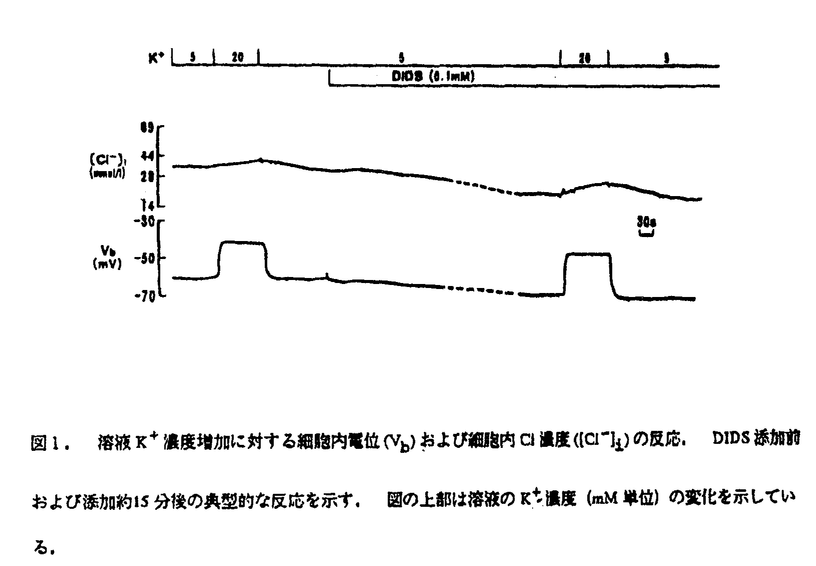 図1.溶液K+濃度増加に対する細胞内電位(Vb)および細胞内Cl濃度([Cl-]i)の反応.DIDS添加前および添加約15分後の典型的な反応を示す.図の上部は溶液のK+濃度(mM単位)の変化を示している. 図1.溶液K+濃度増加に対する細胞内電位(Vb)および細胞内Cl濃度([Cl-]i)の反応.DIDS添加前および添加約15分後の典型的な反応を示す.図の上部は溶液のK+濃度(mM単位)の変化を示している. この[Cl-]iの反応は理論的にa)KおよびClコンダクタンスの共存、b)K-Cl共輸送体、およびc)Na-K-2Cl共輸送体などによると考えられる.これらの可能性を調べるためにまず2種の構造的に異なるKチャンネル阻害剤(バリウムとキニーネ)の作用を検討したが、これらの阻害剤は溶液のk濃度変化に対する脱分極をほぼ完全に抑制すると同時に[Cl-]iの増加反応もほぼ完全に抑制した.従って、[Cl-]iの増加反応は溶液のk濃度変化自体によるものでなく,むしろ細胞内電位の変化によるものであると解釈できる.またClチャンネル阻害剤(A9CとIAA-94)は溶液のk濃度変化に対する脱分極には影響しなかったが、[Cl-]iの増加反応を抑制した.これはa)の可能性(KおよびClコンダクタンスの共存)とよく一致した結果といえる. 一方、K-Cl共輸送体の特異的阻害剤であるH74およびNa-K-2Cl共輸送体の特異的阻害剤であるFurosemideは溶液のk濃度変化に対する反応に影響を与えなかった.これらの結果から[Cl-]iの増加反応は基底側のClコンダクタンスを介する細胞内へのClの流入によることが強く示唆された. このことをさらに確認するために、溶液のCl除去に対する反応を検討したが、Kチャンネル阻害剤の存在下でのみCl除去に対して有意な脱分極が観察された.この起電性の反応はCl電流を反映するものであり、電気的に中性な輸送体であるK-Cl共輸送体およびNa-K-2Cl共輸送体の可能性とは相反するものである.以上の結果から確かに基底側にClコンダクタンスが存在することが示されたが、その大きさはKコンダクタンスに比べかなり小さいことがわかる.従って、過去の報告においてはこの比較的小さなClコンダクタンスが他のより大きなコンダクタンスまたは起電性の輸送体の影響により見過ごされていた可能性が示唆される. 次にこのClコンダクタンスの調節機構についても検討した.上皮細胞のCl channelにはcAMPにより活性化されるタイプのものがあるため、まずForskolinおよびDBcAMPの影響を検討したが、これらの薬剤は溶液のk濃度変化に対する[Cl-]iの増加反応を明らかに増大させた.さらにこの反応はClチャンネル阻害剤(IAA-94)により減弱することから、確かにこのClコンダクタンスが活性化されていることがわかる.一方、Ionomycin(Ca ionophore)およびPMA(PKC acivator)は溶液のk濃度変化に対する[Cl-]iの増加反応を変化させなかった.このことから、この部位のClコンダクタンスはcAMPにより活性化されるが、細胞内Ca濃度の上昇やPKCの活性化には影響されないことが示された.これらの特性は膿胞線雑症(Cystic Fibrosis)の原因遺伝子であるCFTR(Cystic Fibrosis Transmembrane Conductance Regulator)の性質と共通するものであった.実際にCFTRのmRNAがこの部位に存在することも報告されているが、特異的阻害剤がないためこのコンダクタンスがCFTRによるものかどうか現時点では断定することができなかった. 以上の結果から我々は近位尿細管S3部基底側膜に比較的小さなClコンダクタンスが存在し、その活性はcAMPにより増加することを明らかにした.その生理的意義については管腔側膜におけるCl輸送機構が未だ解明されていないため詳細は不明であるが、最近この部位のCl再吸収がcAMPにより増加することが報告されている.さらに、定常状態の細胞内Cl濃度が電気化学的平衡よりも高く保たれていることも、このコンダクタンスが生理的条件下にて近位尿細管Cl再吸収にかなり寄与していることを示唆している.このClコンダクタンスがどのようなタイプのClチャンネルによるものなのかは現時点ではわからないが、これは今後の重要な課題と思われた. |