本研究はヒト由来cDNAライブラリー中から医薬としての可能性を有する分泌蛋白質をコードするものを効率的に選出することを目的として、未知のcDNAとレポーター遺伝子とのキメラ遺伝子を動物細胞で発現させ、その発現産物の局在からこのcDNAが分泌蛋白質をコードしているか否かを判定するアッセイ系の確立を試みたものであり、次の結果を得ている。 1・動物細胞のトランスフェクションの簡素化のため、f1ファージを用いる方法を試みた。SV40初期プロモーター下流に接続されたウロキナーゼ遺伝子またはブラストサイジンS耐性遺伝子を一本鎖DNAの形で含むf1ファージ粒子を、DEAEデキストランまたはリポポリアミン存在下にサル腎由来COS7細胞に加えたところ、これらの遺伝子の発現が確認された。各種ヌクレアーゼ処理した試料についても同様の結果が得られたことから、f1ファージ粒子内の一本鎖DNAがCOS7細胞内で発現することが示された。 2・f1ファージによるトランスフェクションは、ベクターからSV40複製領域を含むSV40初期プロモーターを除去し、かわりにメタロチオネインプロモーターにより遺伝子を発現させた場合にも成立した。このことから、このトランスフェクションにはSV40複製領域は必須ではないことが示された。また、ラージT抗原非産生細胞においてもf1ファージによるトランスフェクションが成立した。 3・f1ファージによるトランスフェクションの至適条件について検討した結果、リポポリアミンの場合、105個のCOS7細胞に対して大腸菌培養液10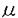 lに相当するファージ粒子により高効率のトランスフェクションが可能であることが示された。 lに相当するファージ粒子により高効率のトランスフェクションが可能であることが示された。 4・未知cDNAの5’端フラグメントにレポーター遺伝子としてウロキナーゼのプロテアーゼドメインcDNAを接続したキメラ遺伝子を動物細胞で発現させ、培地中のレポーター蛋白質の有無からシグナルペプチドの有無を判定するというシステムを考案し、このためのベクターとしてpSSD1を構築した。ウロキナーゼ自身のシグナルペプチドに相当するcDNAの全体あるいは一部をpSSD1に挿入したものの発現を行った結果、完全長ウロキナーゼcDNAと同程度の活性が検出されたのはシグナルペプチドcDNA全体を用いた場合のみであり、シグナルペプチドが不完全な場合は活性は激減または全く検出されなかった。このことから、この方法はシグナルペプチドのコード領域を完全な形で含むcDNAフラグメントのみを特異的に検出できることが示された。 5・既知の分泌蛋白質およびタイプ1膜蛋白質のシグナルペプチドcDNAを用いたモデル実験においては、いずれのキメラ遺伝子についても発現産物のウロキナーゼ活性が培地中に検出され、この方法がウロキナーゼ以外の分泌蛋白質のシグナルペプチドの検出についても有効であることが示された。 6・約600種の新規cDNAの塩基配列からアミノ酸配列を推定し、N末端側に疎水性アミノ酸に富む領域を有する50クローンを選出した。このうち10クローンについて、cDNAの3’末端側からエキソヌクレアーゼを用いて欠失フラグメントを作製し、これらをpSSD1に挿入してキメラ遺伝子を作製した。これらのキメラ遺伝子をCOS7細胞で発現させ、そのウロキナーゼ活性の局在を調べたところ、10種のうち7種では活性が培地中で検出され、シグナルペプチドをコードするものであることが示された。1種では、活性は培地中には全く見出されず、細胞表面において検出された。 7・シグナルペプチドの存在が確認されたクローンの一部についてその全塩基配列を決定し、ホモロジー解析を行ったところ、1クローンがTGF-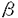 スーパーファミリーの幾つかのメンバーと20〜40%のホモロジーを有していた。さらにC末端側領域中のシスティンの位置、活性化プロセシング部位様配列など、このファミリーの構造上の特徴を示すことから、これに属する新規蛋白質ではないかと考えられた。ノザン解析の結果、このクローンに相当するmRNAは特に胎盤で大量に発現していることが示された。 スーパーファミリーの幾つかのメンバーと20〜40%のホモロジーを有していた。さらにC末端側領域中のシスティンの位置、活性化プロセシング部位様配列など、このファミリーの構造上の特徴を示すことから、これに属する新規蛋白質ではないかと考えられた。ノザン解析の結果、このクローンに相当するmRNAは特に胎盤で大量に発現していることが示された。 以上、本論文ではf1ファージ粒子内の一本鎖DNAが動物細胞内で発現することを初めて明らかにし、これを利用した動物細胞トランスフェクション法の確立について述べた。またシグナルペプチドをコードするcDNAの判定法としてウロキナーゼとのキメラ遺伝子を用いる方法を新たに開発し、実際に新規cDNAに適用してその有効性を証明した。これらの方法は組み合わせることにより、多数の未知cDNA中から分泌蛋白質や膜蛋白質のcDNAを効率的に分離同定する上で有用である。本研究は医薬として有用な蛋白質の探索、開発に貢献をなすと考えられ、学位の授与に値するものと考えられる。 |