| 内容要旨 | | 緒言 メタン資化細菌はメタンを唯一の炭素源かつエネルギー源として増殖するという独特の生理的性状に基づいてまとめられた細菌のグループである。メタン資化細菌は次の3つの点で近年注目されている。1.メタンは二酸化炭素に次ぐ地球温暖化作用をもつ気体であるが,メタン資化細菌は大気メタン濃度増加に対して抑制的に働く。2.大洋底の沈み込み域に存在するメタン湧出域では,メタン資化細菌を鰓に共生させる後生動物が知られており,メタン資化細菌はこれらの栄養源となっている。3.メタン資化細菌は通常では分解されにくい有機ハロゲン化物を分解するため,土壌中に蓄積されているこれらの物質の分解に役立つ可能性がある。 これらのことから,海洋のメタン資化細菌の分布,分類及び機能について関心が持たれるが,海洋に於けるメタン資化細菌の研究は1949年のHutton and ZoBellによる最初の分離からしばらく研究がなく,80年代の終わりから現在にかけていくつかの分離の報告があるのみである。また,現存している海産メタン資化細菌株の培養株は数株にすぎない。そこで,本研究では沿岸環境中のメタン資化細菌の存在を明らかにする目的で,計数を試みた。また,沿岸海底泥から純粋分離を行い,同定するとともに生理的性状について検討した。とくに,海産メタン資化細菌のメタン利用速度と増殖との関係はメタン資化細菌の存在と活性とを関連づけるのに必要と考えられるので培養系において検討した。 MPN法によるメタン資化細菌の計数 計数に先立ち,MPN法に用いる培養の条件を設定し,得られる増殖がメタン資化細菌であるかどうかを検討した。沿岸海底泥1mlを完全合成無機液体培地(B728)100mlに接種し,内容量1.5リットルの培養容器の気層の1/2をメタンで置換し,35℃振盪及び20℃静置で培養を行った。1〜6週間のうちに,茶色,ピンク色,乳白色などの着色があるフロックまたは菌膜状になる増殖が得られた。得られた培養はメタンを添加しないとタンパク質の増加はなく,メタンの存在によってのみタンパク質の増加がみられた。また,メタンと酸素を消費し,二酸化炭素を放出して増殖することが確認された。さらに,これらのうち菌体の透過電顕切片像による内部形態の観察を行った3つの培養にはメタン資化細菌に特徴的な内膜構造を持つ細胞が観察された。 続いて同様の条件で海底泥をインキュベートし,メタン資化細菌のMPN法による計数を試みた。その結果,東京湾及び大槌湾の底泥サンプル中には最高2.4×103cells/ml sediment分布していた。得られた計数値の範囲はほぼ101〜103であり,これはHeyer et al.(1984)による黒海における計数と同程度の範囲であった。計数方法の改良として,メタンの減少と同時に酸素の減少と二酸化炭素の増加を検出することによりメタン資化細菌増殖を確認する方法を検討した。その結果,菌膜を形成したものを陽性とした場合とメタンと酸素の減少と二酸化炭素の増加があったものを陽性にした場合とでは,後者の方が計数値が高くなるか等しいかのどちらかであった。この原因は,菌膜の有無の肉眼的判断よりもガスクロによるガスの変化の方が検出感度がより高いことによると考えられる。 海底泥中のメタン濃度の鉛直プロファイルとメタン資化細菌の分布との間には,東京湾のメタンが検出された底泥では,メタン資化細菌の計数値が高いところでメタン量が減少するという関係が観察され,底泥から水中へのメタンの放出をメタン資化細菌が抑制している可能性が示唆された。 メタン資化細菌の分離・同定と生理的性状 海産に限らずメタン資化細菌は純粋になりにくいものが多いことが従来から知られていた。本研究に於いても,B728培地を1.2%の寒天で固化させた平板培地上のコロニー形成による集積培養からの純化を試みたが,コロニーはきわめて高濃度に接種したときにのみ形成され,純化はできなかった。そこで,純粋分離させるべく試したいくつかの別の方法のうち,通常用いられる寒天平板培地の1/3〜1/4の濃度である0.4%程度のアガロースゲルで緩やかに固化させた無機完全合成培地IM-310を用いることでメタン資化細菌のコロニーを形成させることが可能となった。IM-310培地はB728培地がリン酸塩によってpH緩衝していたのを炭酸塩に入れ替える等の変更を加えたものである。 この方法によって,大槌湾底泥試料から4株,東京湾底泥試料から3株を純粋分離し,この7株について,性状試験と併せて,リン脂質脂肪酸(PLFA)の組成及び16SrRNAの部分塩基配列の分析を行い同定した。 Bowman et al.(1993)は,124項目の性状によって数値分類を行ってメタン資化細菌を16のPhenonに分けた。これらの中で海産のものが含まれるのはphenon5とphenon9のみであった。彼らの調べた海産メタン資化細菌は全て増殖にNaClを要求するという共通点があった。phenon5とphenon9の2群は,他のいくつかのphenonと併せてMethylobacter属に分類されている。 本研究で純粋分離した7株は,全て増殖にNaClを要求し,3.0%のNaCl濃度でも増殖した。7株うち,O-12a株以外は5.0%のNaCl濃度でも増殖が可能であった。7株のPLFA分析及び性状試験の結果,大槌湾産の株はMethylobacter属であり,phenon5と考えられた。また,東京湾産の株はMethylococcus属であり,phenon11であると考えられた。Methylococcus属の海産メタン資化細菌はこれが初めての分離である。16SrRNAの部分塩基配列を用いて描いた系統樹も,大槌湾産のものと,東京湾産のものとは,離れたそれぞれのグループに分かれた。Phenon9には海産種しか含まれていないが,16SrRNAの部分塩基配列からはphenon5とphenon11の中間的な位置を占めた。 海産メタン資化細菌の増殖とメタン消費 大槌湾から分離した株のO-12a及びO-4a,東京湾から分離した株の07061及び9232,並びに陸環境から分離されたMethylobacter albus(NCIMB 11123)を以後の実験に供した。培養実験は,500mlの耐圧濾過瓶に吸光度測定用の試験管とガスサンプル採取用のセプタムを取り付けた内容量810mlの培養器を用いた回分培養で行った。培地はIM-310培地50mlを用い,培養器内1atmのうち,メタンと空気を11:89となるようメタンを添加し,往復振盪培養行った。前培養は実験条件と同じ条件で培養し,660nmにおける吸光度(A660)が計算上0.001となるように新たな培地に添加し,ガス置換によって培養を開始した。ほぼ一定時間間隔でA660の測定を行うとともにガスクロマトグラフによってメタン及び酸素,二酸化炭素の定量を行った。 それぞれの株について様々な培養条件で得られた増殖曲線から,それぞれの最大比増殖速度( max)を求めた。maxはO-4a及びO-12aでは,20℃から30℃にわたってほぼ一定であり,それ以下では低下し,35℃では増殖しなかった。また07061及び9232は,35℃にピークがあり,それ以下の温度ではなだらかに低下した。さらに,9232は35℃以上では急激に低下し,07061は40℃では増殖しなかった。Methylobacter albusは35℃にピークがあり,それ以下では減少し,20℃では増殖しなかった。IM-310培地のNaCl濃度を変化させてmaxへの影響を調べた。5つの株とも,0.001Mまたは,0.01Mで高いmaxの値が得られ,高塩分ではmaxは低下した。しかしながら,海産の4株は,全て0.7MNaClでも増殖したのに対し,Methylobacter albusは0.25M NaCl以上では増殖せず,他の海産の株とは異なっていた。今回対象とした海産メタン資化細菌では,15℃〜30度で得られる平均的maxは,0.07h-1であった。 max)を求めた。maxはO-4a及びO-12aでは,20℃から30℃にわたってほぼ一定であり,それ以下では低下し,35℃では増殖しなかった。また07061及び9232は,35℃にピークがあり,それ以下の温度ではなだらかに低下した。さらに,9232は35℃以上では急激に低下し,07061は40℃では増殖しなかった。Methylobacter albusは35℃にピークがあり,それ以下では減少し,20℃では増殖しなかった。IM-310培地のNaCl濃度を変化させてmaxへの影響を調べた。5つの株とも,0.001Mまたは,0.01Mで高いmaxの値が得られ,高塩分ではmaxは低下した。しかしながら,海産の4株は,全て0.7MNaClでも増殖したのに対し,Methylobacter albusは0.25M NaCl以上では増殖せず,他の海産の株とは異なっていた。今回対象とした海産メタン資化細菌では,15℃〜30度で得られる平均的maxは,0.07h-1であった。 メタン資化細菌はメタンのみを有機物源とし,好気的に増殖する。窒素源として充分な硝酸態窒素を与えた場合,メタン資化細菌の増殖は次のように考えられる。  ただし,右辺の第1項はメタン資化細菌のバイオマスに相当する。 また,基質量の消費速度を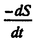 ,細菌の増殖速度を ,細菌の増殖速度を ,細菌バイオマスをM,としそれぞれ炭素量で表し,炭素量で表したバイオマス収率をyc*,細胞の維持に用いられる細菌炭素あたりの基質利用速度をmcとすると, ,細菌バイオマスをM,としそれぞれ炭素量で表し,炭素量で表したバイオマス収率をyc*,細胞の維持に用いられる細菌炭素あたりの基質利用速度をmcとすると,  が成立する。メタン資化細菌の増殖と,メタン消費速度をこのモデルに当てはめ,yc*,mcを求めた。これらの値は,株または培養条件による大きな変動は見られず,平均的なyc*の値として0.525が,mcは0.00658がそれぞれ得られた。また,yc*からエネルギー効率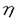 *を求めると,0.282となった。yc*の値は,既に報告されているメタン資化細菌についての値(0.318〜0.616)の範囲の中で,やや高めの値であった。 *を求めると,0.282となった。yc*の値は,既に報告されているメタン資化細菌についての値(0.318〜0.616)の範囲の中で,やや高めの値であった。 以上の結果で得られた数値から,次の式によって,メタン資化細菌のバイオマスからメタン消費速度を推定することができる。 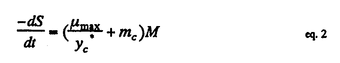 環境中の値として報告されているメタン資化細菌数とメタン消費速度の値との両対数値の回帰直線の勾配は0.78(r2=0.88)であったが,1と有意差は認められず,傾きを1とみなしたときも高い寄与率(r2=0.72)が得られた。このことは,eq.2のメタン資化細菌のバイオマスとメタン消費速度との関係を用いて,環境中に於いてもメタン資化細菌のバイオマスからメタン消費速度を推定可能であることを意味している。バイオマスが炭素量で与えられるときにはeq.2の括弧の中は本研究で得られた値から0.043となる。バイオマスが細胞数で与えられるときには,本研究で得られた海産メタン資化細菌についての値230fgC/cellを用いて炭素量に換算することができる。これによって得られた推定値は,環境中のメタン消費速度の値に対して高い寄与率(r2=0.62)であった。 |