| 1.はじめに 酸素発生型の光合成生物に含まれるクロロフィル(Chl.)には、Chl.a、b、c(c1、c2)がよく知られている。Chl.d(図1)は、微量な緑色色素として、さまざまな大型紅藻から検出された(Manning & Strain,1943)。しかし、この色素を確実に含む藻類が確定されていないこと、また、Chl.aの酸化により生成され得ることなどから、Chl.dは色素抽出過程における人工産物であり天然に存在しないという考え方が近年の主流であった。  図1 クロロフィルdの化学構造クロロフィルaは矢印の部分が -CH=CH2 図1 クロロフィルdの化学構造クロロフィルaは矢印の部分が -CH=CH2 藻類における光合成色素の多様性を調べる目的で、パラオ諸島沿岸の群体ホヤから微細藻類を分離した。群体ホヤ、特にディデムニ科のは、Prochloronをはじめ特殊な色素系をもつ藻類を共生させていることで知られている原索動物である。その結果、Chl.dを主要色素としてもつ原核光合成生物Acaryochloris marina Miyashita et Chihara gen.et sp.nov.が分離された。 本論文では、A.marinaの形態について述べ、吸収スペクトル測定および色素組成分析結果からChl.dが天然に存在することを示した。また、光合成酸素発生測定によりChl.aを主要色素としない光合成系の存在を明かにした。さらに、他の光合成生物との分子系統関係を調べ、A.marinaおよびクロロフィルdの起源について考察し、A.marinaの分類学的位置の検討を行った。 2.生物学的特徴 パラオにて採取した群体ホヤの中心部をナイフで切断して、体内の藻類をピペットで吸い出し、海水をベースとするK+ESM培地(Miyashita et al.,1995)に植えて集積培養した。培養2ヶ月後、生育した藻類を寒天プレート上で単藻化し、A.marinaを単離した。培養は、K+ESM培地中で、28℃、空気通気、50-100 mol photons m-2s-1の白色蛍光灯を照射(12時間/日)して行った。 mol photons m-2s-1の白色蛍光灯を照射(12時間/日)して行った。 A.marinaは、直径1.0-2.0mの球形から楕円形の単細胞原核生物であった(図2)。染色体やリボゾーム、カルボキシソームは細胞中心部に散らばって存在していた。チラコイド膜は細胞の内周に沿って互いに密に詰まった状態で存在していた。チラコイド膜上にフィコビリゾームは認められなかった。A.marinaのチラコイド膜構造は、ラン藻類よりむしろ原核緑色藻類(Prochlorophyta)に似ていた。 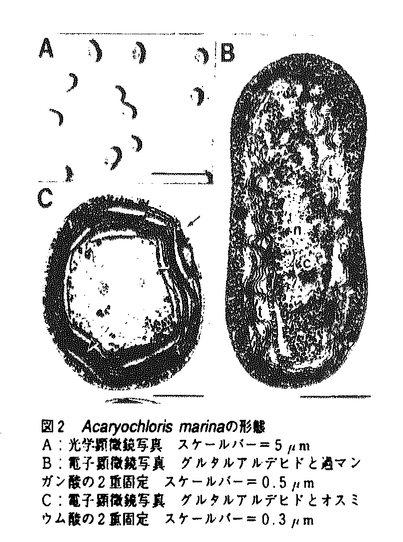 図2 Acaryochloris marinaの形態A:光学顕微鏡写真 スケールバー=5m B:電子顕微鏡写真 グルタルアルデヒドと過マンガン酸の2重固定 スケールバー=0.5m C:電子顕微鏡写真 グルタルアルデヒドとオスミウム酸の2重固定 スケールバー=0.3m 図2 Acaryochloris marinaの形態A:光学顕微鏡写真 スケールバー=5m B:電子顕微鏡写真 グルタルアルデヒドと過マンガン酸の2重固定 スケールバー=0.5m C:電子顕微鏡写真 グルタルアルデヒドとオスミウム酸の2重固定 スケールバー=0.3m A.marina細胞は緑色で、716nm付近に吸収極大を示した(図3)。既知の酸素発生型の光合成生物の吸収極大(680nm付近)に比べ、30nm以上長波長であった。そこで細胞に含まれる色素をメタノールで抽出し、赤色光域(650-750nm)の吸収極大を測定したところ(図4)、Chl.aやbの吸収(図5)と異なる697nmに吸収極大を示したことから、A.marinaの主要色素が特異な色素であることが示された。  図3 細胞懸濁液吸収スペクトルの比較 図3 細胞懸濁液吸収スペクトルの比較 図4 細胞のメタノール抽出液の吸収スペクトル 図4 細胞のメタノール抽出液の吸収スペクトル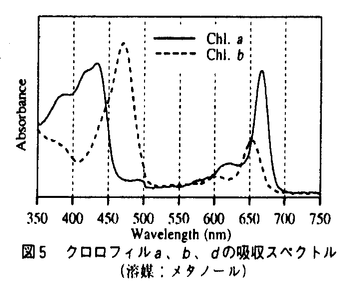 図5 クロロフィルa、b、dの吸収スペクトル(溶媒:メタノール) 図5 クロロフィルa、b、dの吸収スペクトル(溶媒:メタノール) さらにメタノール抽出液中の色素をHPLCにより分離したところ、溶出順にMgDVP、ゼアキサンチン、Chl.d、Chl.a、 -カロテンの5色素が検出された(図6)。各色素はHPLC分析における保持時間と色素の吸収スペクトルを標準色素と比較して同定された。しかし、Chl.dの標準色素が存在しなかっため、ピーク3の色素については、精製後、各種溶媒中での吸収ピークの比較、およびNMRによる構造決定の後に、Chl.dと同定された。 -カロテンの5色素が検出された(図6)。各色素はHPLC分析における保持時間と色素の吸収スペクトルを標準色素と比較して同定された。しかし、Chl.dの標準色素が存在しなかっため、ピーク3の色素については、精製後、各種溶媒中での吸収ピークの比較、およびNMRによる構造決定の後に、Chl.dと同定された。  図6 色素のHPLCクロマトグラム 図6 色素のHPLCクロマトグラム HPLCチャートのピーク面積と各色素の吸光係数(440nm)に基づいて色素含量をを算出したこところ、Chl.a含量は1mg/g dry cells未満であった(表1)。既知の光合成生物のChl.a含量(3〜30mg/g dry cells)に比べはるかに小さく、A.marinaはChl.a含量の最も低い光合成生物であった。これに対し、Chl.dは20mg/g dry cells以上含まれていた。ゼアキサンチン、-カロテンはそれぞれ2mg/g dry cells程度含まれていた。この結果から、A.marinaの主要色素がChl.dであることが確認された。  表1 A.marinaのもつ色素とその吸収ピークおよび含量注:Peak No.は図6のピークに対応する。Amaxは、HPLCで溶出した色素の吸収極大。 表1 A.marinaのもつ色素とその吸収ピークおよび含量注:Peak No.は図6のピークに対応する。Amaxは、HPLCで溶出した色素の吸収極大。 透過型電子顕微鏡観察ではフィコビリゾームが認められなかったが、細胞を破砕したときの水溶性画分にフィコビリンタンパク質(フィコシアニン、アロフィコシアニン)が認められ、A.marinaのフィコビリンタンパク質の存在形態が、ラン藻類や紅藻類のものと異なることを示唆していた。 Chl.dを主要色素とするA.marinaは、光照射下、光合成酸素発生を行った(図7)。酸素発生は光強度の増加とともに増加し、200mol photons m-2s-1付近で飽和した。これまで酸素発生型の光合成には、Chl.aを主要色素とすることが不可欠であると信じられてきた。これはChl.aが光エネルギーの捕獲、伝達および初期電荷分離に最も重要な役割をになっているためである。しかし、A.marinaが酸素発生型の光合成を行うことから、酸素発生型の光合成には、必ずしもChl.aを主要色素とする必要のないことが明かとなった。  図7 光合成酸素発生3.分子系統関係 図7 光合成酸素発生3.分子系統関係 A.marinaがどのような生物に最も近縁か、またどのような系統において出現した生物か知見を得るため、rbcLのアミノ酸配列ならびに16SrDNA遺伝子の塩基配列を決定し、既知の光合成原核生物との分子系統関係について調べた(図8)。その結果、両系統解析結果において、明らかにA.marinaがラン藻類に最も近縁であると同時に、ラン藻類の系統群の中に位置することがわかった。つまりChl.dを主要光合成色素とするという極めて特異な色素組成をもつA.marinaが、ラン藻類や原核緑色藻類から独立した生物ではなく、むしろそれらと同じ祖先生物から派生した生物であることが示唆された。さらにChl.dを合成し、これを光合成色素として使用するという光合成のメカニズムは、A.marinaもしくはA.marinaの祖先生物において独自に獲得されたものであると考えられた。紅藻類中におけるChl.dの存在については、いまだ不確定である。仮に紅藻類がChl.dをもつ場合、その色素組成は、Chl.aとdおよびフィコビリンタンパク質を含むという点においてA.marinaと一致する。しかし系統樹は、A.marinaと紅藻類の葉緑体に直接的な系統関係が存在しないことを示した。つまりChl.dが紅藻類に含まれるとしても、A.marinaにおけるChl.dの獲得は、紅藻類におけるChl.dの獲得とは異なった系統において、独立して起こったものであると考えられた。  図8 16SrDNA配列を用いた最大節約法による分子系統樹の一例4.分類学的考察 図8 16SrDNA配列を用いた最大節約法による分子系統樹の一例4.分類学的考察 A.marinaは、オルガネラをもたない酸素発生型の光合成原核生物である。原核藻類は現在、主要光合成色素の違いからChl.aとフィコビリンタンパク質をもつことを特徴とするラン色植物門と、Chl.aとbをもちフィコビリンタンパク質をもたないことを特徴とする原核緑色植物門に分けられている。この分類に従えば、Chl.dを主要光合成色素としChl.aとフィコビリンを微量含有するA.marinaには、新たな門の提唱が必要となる。しかし、本研究において得られた系統関係では、A.marinaが他の酸素発生型光合成原核生物から独立した生物ではないことが示され、新門の提唱は分子系統関係上支持されなかった。つまり、光合成色素を主な指標としている原核藻類の分類方法が分子系統上支持されないことがわかった。そこで植物分類群において、ラン色植物門Cyanophytaならびに原核緑色植物門Prochlorophytaをそれぞれ綱に格下げし、原核藻類全体を定義する門として、原核植物門(Prokaryophyta)を提唱した。A.marinaは、原核植物門のなかに新綱Acaryochloridophyceae、新目Acaryochloridales、新科Acaryochloridaceaeを設立し、ここに帰属された。 5.おわりに A.marinaの発見により、Chl.dが天然に存在することが証明された。また、酸素発生型の光合成には、必ずしもChl.aを主要色素とする必要がないことが初めて明かにされた。A.marinaには新たな光合成メカニズムの存在が期待される。さらに分子系統解析結果から、A.marinaがラン藻類や原核緑色藻類と同じ祖先生物から出現したことが示唆された。 A.marinaの存在は、原核藻類が進化の過程において既存の生物からは想像もできない光合成色素系の多様性を獲得していることを示している。今後も特異な光合成色素系をもった藻類の発見が予想され、光合成色素系の進化、光合成生物進化の解明に大きな役割を果たすものと期待される。本研究をその多様性を明らかにする第一歩と位置づけている。 |