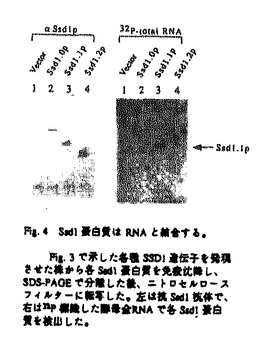
本論文は2章からなり、第1章ではSsd1蛋白質がRNAと結合する蛋白質であることを生化学的に証明した。第2章では、Ssd1蛋白質と結合する蛋白質を2-hybrid法で検索し,1つの遺伝子産物Fis1を得た。Fis1蛋白質は既にmRNAの3’末端のプロセッシングに関与する因子として報告されていた蛋白質と同一であることが分かった。本論文では、これらの情報を総合してSsd1-Fis1蛋白質のmRNAの代謝にかかわる作用についてモデルを構築している。以下に本論文の内容を要約する。 酵母Saccharomyces cerevisiaeのSSD1遺伝子は、protein phosphatase、MAPKK kinase、RNA polymerase IIIのサブユニット、cAMP phosphodiesteraseをそれぞれコードするSIT4、BCK1、RPC31、PDE2遺伝子の変異を1コピーで抑圧する遺伝子として単離された。また、菊池らによりミニ染色体の安定保持に欠損を示すmcs1変異を1コピーで抑圧する遺伝子としても単離されている。さらにG1サイクリンをコードするCLN1、CLN2遺伝子の二重変異株とssd1変異の三重変異株は合成致死となり、MAP kinaseをコードするMPK1遺伝子とssd1変異の二重変異は高温で増殖不能となる。これらの事実は、SSD1が多種類の制御系に関与する興味深い遺伝子であることを遺伝学的に示している。この多様な遺伝的相互関係から様々な機能が類推されていたが、実際の機能は全く不明であった。本研究はSSD1遺伝子の機能を解明することを目的とし、解析を行ったものである。 第1章 Ssd1蛋白質の生化学的機能解析1)SSD1-v及びssd1-d遺伝子の生化学的性格付け Amdtらの遺伝学的解析により、SSD1はviable type(SSD1-v)とdead type(ssd1-d)、二種の遺伝子型で存在することが予想されていたが、抗体を用いた解析より、SSD1-vは約160kD、ssd1-dは120kDの蛋白質を各々コードしている事実を生化学的に明らかにした。またW303株だけでなくYPH499など酵母研究で野生型基準株として使用される株中でもssd1-dとして存在していた。この事実はSSD1遺伝子のdead typeが、研究用酵母株で広く蔓延していることを意味する。従ってdead typeと任意の変異との二重変異が相加的な表現形を示すケースではSSD1遺伝子が1コピー抑圧遺伝子として単離されると考えられる。 2)Ssd1蛋白質の機能解析 Ssd1蛋白質には、出芽酵母のSpDis3ホモログScDis3、ミトコンドリアに局在するDss1、分裂酵母の細胞周期の進行に関わるSpDis3、細胞形態の維持に関わるSts5、アカパンカビのミトコンドリアに局在しmtRNAの修飾に関与するCyt4、赤痢菌の毒素発現に転写後レベルで関与するVacB、さらに大腸菌の3’-5’exoribonucleaseをコードするRnbなどの蛋白質と種を超えて相同性を示す領域が存在する。しかしながら、これらの蛋白質の機能はRnbを除いて明らかになっていない。 ssd1遺伝子破壊株は高温で増殖不能となるが、完全長のSSD1遺伝子を導入すると増殖は回復するのに対して、最も保存された領域(D3)を欠くSSD1.1を導入しても回復しない。従って、D3はSSD1遺伝子の機能発現に必要であると考えられる。D3を持つ、異種間のSsd1ホモログ遺伝子のなかで重要な共通項はRNAの修飾に関与する可能性である。まず免疫沈降により精製したSsd1蛋白質を用いて、放射性標識したpoly(rA)を基質としexoribonucleaseの活性を測定したが活性は見られなかった。そこで次にSsd1蛋白質がRNAと結合する可能性を検討した。放射性標識した酵母全RNAをプローブとしてNorth-Western法を行った結果、Ssd1蛋白質はRNAと結合する事実が明らかになった。また各種のpoly nucleotideをリガンドとしたアフィニティー精製実験により、Ssd1蛋白質はsingle strand DNAに効率よく結合し、poly(rA)に最も親和性の高いことがわかった。これらの事実は共通領域を持つSsd1以外の機能不明の蛋白質の解明に貢献すると考えられる。またSsd1蛋白質はリン酸化されているので、その機能は蛋白質キナーゼにより制御を受けていると考えられる。 Ssd1蛋白質は主に細胞質に存在していることから、核内ではなく細胞質側のRNAと相互作用していると考えられる。さらに大部分のSsd1蛋白質は細胞質内のポリソーム画分には存在していないため、Ssd1は活発に翻訳されている状態のmRNAとは相互作用していないと考えられる。これに対して分裂酵母Dis3の機能ホモログである出芽酵母のScDis3は核内に、Dss1はミトコンドリアに局在する可能性が示されている。出芽酵母で上述の共通領域を持つ蛋白質はこれら以外にはない。従って、Ssd1、ScDis3、Dss1蛋白質は細胞質、核、ミトコンドリアのRNAにそれぞれ作用している可能性が考えられる。 第2章 Ssd1蛋白質と結合する因子の解析 Ssd1蛋白質の細胞内での役割をさらに詳細に解析するため、Ssd1蛋白質と結合する因子をTwo-hybrid法により検索した。その結果、第五番染色体のYER032遺伝子を単離し、FIS1(Factor Interacting with Ssd1)と名付けた。FIS1遺伝子にはセリン、スレオニンに富む925個のアミノ酸をコードしうる一つのORFが存在し、遺伝子破壊の結果、増殖には必須でないことが分かった。Ssd1蛋白質とFis1蛋白質が実際に細胞内で結合していることは、抗体を用いた免疫沈降実験により確かめた。 Fis1蛋白質は核内のmRNA生成過程での3’側切断に必要とされるRef2の結合因子Fir1、またmRNAのpoly(A)付加酵素Pap1の結合因子Pip1と同一であることが明らかになったため、Ssd1、Fis1/Fir1/Pip1、Ref2、Pap1の四者は複合体を形成し、様々なRNAのなかでも特にmRNAに作用している可能性が示唆された。しかしながら、Ssd1蛋白質は主に細胞質に存在しているのに対して、Ref2、Pap1は核に存在すると考えられている。この事実は四者が単純に一つの複合体を形成しているのではないことを示唆している。そこでFis1-Ref2、Fis1-Pap1とFis1-Ssd1が別々の複合体を形成して核と細胞質にそれぞれ存在すると仮定し、さらにmRNAの3’末端を修飾する他の遺伝子産物群とFir1、Ssd1の相互関係をTwo-hybrid法により検討した。その結果、Fis1蛋白質がSsd1、Ref2だけでなく、核、細胞質両方に存在するpoly(A)結合蛋白質Pab1、細胞質に存在するpoly(A)分解酵素Pan3とも結合しうることを明らかにした。さらにこれらの因子が結合するFis1の領域をTwo-hybrid法により検索したところ、いずれの結合においてもFis1のC末端側の約200アミノ酸の領域を必要とすることが明らかになった。この事実はこれらの因子が巨大な複合体を形成しているのではなく、Fis1との各々別の複合体を形成していることを示唆している。従ってFis1-Ref2、Fis1-Pap1複合体は核内、Fis1-Pab1は核、細胞質、またFis1-Pan3、Fis1-Ssd1複合体は細胞質でmRNAの3’末端に作用する可能性が考えられる。 本研究により、リン酸化蛋白質Ssd1はRNAに結合することが明らかになった。またSsd1結合蛋白質Fis1はmRNAの3’側の修飾に関与する蛋白質群と物理的に相互作用するので、Ssd1もmRNAの3’側の修飾反応に関わる可能性が考えられる。現在、出芽酵母mRNAの分解制御機構は5’側からの機構が詳細に研究されているが、3’側からの分解制御機構はほとんど明らかにされていない。またmRNAのpoly(A)に結合する因子は翻訳開始制御と密接な関係をもつため、3’末端を介したmRNAの安定性制御機構の解明は重要であると考えられる。本研究の成果はその詳細な理解に貢献するものである。 以上のように、本研究で得られた結果は多くの新しい知見を含み、これらの研究を通じ提出者が分子生物学の分野の学識を十分に持つものであることが確認された。本研究のある部分は東江昭夫氏らとの共同研究であるが、いずれの場合も提出者が中心となって研究を進めたことは明らかであった。以上を総合して、提出者が博士(理学)の学位の授与に値するものであることが確認された。 | 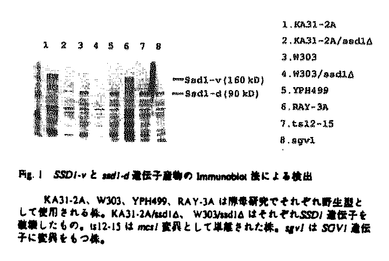
 、W303/ssd1
、W303/ssd1