学位論文要旨
| No | 213569 | |
| 著者(漢字) | 三島,一彦 | |
| 著者(英字) | ||
| 著者(カナ) | ミシマ,カズヒコ | |
| 標題(和) | 悪性グリオーマ細胞におけるヘパリン結合性epidermal growth factor(EGF)Familyタンパクの発現と機能解析 | |
| 標題(洋) | ||
| 報告番号 | 213569 | |
| 報告番号 | 乙13569 | |
| 学位授与日 | 1997.10.22 | |
| 学位種別 | 論文博士 | |
| 学位種類 | 博士(医学) | |
| 学位記番号 | 第13569号 | |
| 研究科 | ||
| 専攻 | ||
| 論文審査委員 | ||
| 内容要旨 | グリオーマ細胞の増殖や悪性化には、epidremal growth factor(EGF)/EGF receptor(EGFR)システムを介するシグナル伝達機構が重要な役割を果たしていると考えられている。EGFRを活性化するリガンドとして、transforming growth factor 本研究に先立ち私は、EGFファミリーに属する増殖因子の中でも、ヘパリンに結合性を有するSDGFが、オートクライン増殖因子としてラットのグリオーマ細胞の増殖に深く関与していることを報告した(Mishima et al.,Int.J.Cancer,66:352-357,1996)。ところが、構造的にSDGFのhuman counterpartと考えられているamphiregulin(AR)の発現は、悪性グリオーマ組織で検出し得なかった。この結果より、ヒトではAR以外のヘパリン結合性EGFファミリータンパクが、グリオーマの細胞増殖、造腫瘍性に関与している可能性があると考えられた。そこで、ヘパリン結合性EGFファミリーのもう一つの代表的な因子であるHB-EGFについて、ヒト悪性グリオーマにおける発現を検討し、さらにその機能解析へと研究を進めた。 HB-EGFは、U-937細胞の培養濾液から、血管平滑筋細胞に対する増殖活性を指標に液性因子として東山らにより分離、精製された。HB-EGFのcDNAがクローニングされ、その構造解析が行われた結果、膜結合タンパク(proHB-EGF)として合成され、細胞外部分がプロセッシングを受け、分泌型となることが判明した。分泌型のHB-EGFは、平滑筋細胞、ケラチノサイト、肝細胞に対して増殖活性を示し、平滑筋細胞に対しては、遊走因子としても働くことが示された。さらに、平滑筋細胞に対するHB-EGFの増殖活性や遊走因子としての活性は、EGFやTGF 本研究では、このように他のEGFファミリーに属するタンパクとは活性や機能の面で異なるHB-EGFについて、悪性ヒトグリオーマ細胞及び、悪性グリオーマ組織における発現を検討し、その機能解析を行った。 HB-EGFcDNAをプローブとしたNorther blot解析の結果、検討した7種類全ての悪性ヒトグリオーマ細胞株においてHB-EGFmRNAの発現が認められた。また、ヒト悪性グリオーマ(glioblastoma)組織では非腫瘍部脳に比較し、2倍から5倍のHB-EGFmRNAの発現を認めたが、ARmRNAは非腫瘍部脳、腫瘍組織のいずれにおいても検出できなかった。 グリオーマ細胞の細胞膜をビオチン化し、膜結合型proHB-EGFを特異的に認識する抗体を用いて免疫沈降した後、HB-EGFタンパクの発現をWestem blot法を用いて解析したところ、約30kDaの膜結合型proHB-EGFに相当するバンドが認められた。以上より、グリオーマ細胞は、細胞表面に膜結合型proHB-EGFタンパクを発現していることが明らかとなった。 悪性グリオーマ組織でのHB-EGFの発現を確認するために、そのレセプターとして知られているEGFRの発現とを併せて、それぞれの特異的抗体を用いた免疫組織化学染色法で検討した。抗EGFR抗体では、腫瘍細胞が染色され、血管内皮細胞は染色されなかった。抗proHB-EGF抗体では腫瘍細胞と、血管増生の部位での血管内皮細胞、及びpericyteと思われる細胞が陽性像を示した。34例の悪性グリオーマ(anaplastic astrocytoma9例、glioblastoma25例)組織中、EGFRが陽性であったanaplastic astrocytomaの3例中2例(66%)で、また、glioblastomaの13例では全例(100%)でproHB-EGFも陽性であった。さらに、EGFR陰性のanaplastic astrocytomaの2例、glioblastomaの4例においても、proHB-EGFは陽性であった。 proHB-EGFは、ジフテリア毒素(DT)のreceptorとしても機能していることが示されている。そこで、グリオーマ細胞で発現が確認されたproHB-EGFがDTのreceptorとして機能し、グリオーマ細胞がDT感受性を示すかをタンパク合成阻害能を指標に検討した。グリオーマ細胞では、10ng/mlのDT投与によって、DT未投与のコントロール細胞でのタンパク合成の約20%まで阻害されることがわった。また、proHB-EGFタンパクを発現していないmouse L cellでは、DTによるタンパク合成阻害能は認められなかった。以上の結果は、proHB-EGFがDT receptorとしても機能しうるというこれまでの報告に矛盾しないものであった。 グリオーマ細胞に対し、HB-EGFは増殖活性を有し、またオートクライン増殖因子として機能しうるかを検討した。血清除去下で培養したグリオーマ細胞に、種々の濃度でrecombinant HB-EGF(rHB-EGF)を投与した結果、濃度依存的に細胞増殖増強効果が認められ、10ng/mlのrHB-EGF投与では、未投与のコントロール細胞に比較し、細胞数が約2倍に増加することがわかった。一方、HB-EGFに対する特異的な中和抗体を加えると、コントロールのIgGを加えたものに比較し、約30-40%の細胞増殖抑制が認められた。以上の結果より、HB-EGFはグリオーマ細胞に対して増殖活性を有し、オートクライン増殖因子として機能していることが明らかとなった。 basic FGF(bFGF)及びEGFは、グリオーマ細胞に対して増殖因子として働くことが示されている。そこで、これらのグリオーマに対する増殖活性をDNA合成能を指標にHB-EGFのそれと比較した。10ng/mlのrHB-EGFの投与は、5%血清投与に比べると、約1.2-2.3倍程度、グリオーマ細胞でのDNA合成能を促進することがわかった。このDNA合成促進能は、10ng/mlのEGF,bFGFに比較すると、10-20%とわずかながら高い値であった。 悪性グリオーマ細胞におけるHB-EGFの発現が、HB-EGF自身、あるいは他の増殖因子による発現制御を受けるかを、T98G細胞を用いて検討した。rHB-EGF投与により、またEGF,TGF 血管内皮特異的な増殖因子であるvascular endothelial growth factor(VEGF)は、グリオーマの血管新生に重要な役割を果たしていることが示されている。HB-EGFは、直接内皮細胞に対して増殖活性をもたないが、VEGFを誘導する能力があれば間接的に血管新生にも関与しうると考えた。そこで、rHB-EGFをグリオーマ細胞に投与し、VEGFが誘導されてくるかを検討した。10ng/mlのrHB-EGFを投与することで、今回検討した3つのグリオーマ細胞株のいずれにおいてもVEGFmRNAの発現が亢進することがわかった。また、このVEGFmRNAの発現誘導は、加えるrHB-EGF濃度に依存して増加することが示された。さらにrHB-EGFによるVEGFmRNAの発現誘導が、VEGFタンパクの培養液中への分泌促進に結びつくことが、VEGF抗体を用いたWestem blot解析の結果から確認された。 本研究より、HB-EGFはオートクライン増殖因子としてグリオーマ細胞の増殖に深く関与していることが明らかとなった。免疫組織化学染色の結果から、proHB-EGFタンパクは悪性グリオーマ組織でも発現しており、かつ、そのレセプターであるEGFRと共発現している腫瘍が多く存在することから、in vivoにおいてはオートクライン機構のみではなく、ジャクスタクライン機構を介しても、EGFRを発現する近接細胞に増殖シグナルを伝達していることが示唆された。グリオーマ細胞での産生が報告されているTGF | |
| 審査要旨 | 本研究は、ヒトグリオーマ細胞の増殖と悪性化に重要な役割を果たしていると考えられている、epidermal growth factor receptor(EGFR)を介するシグナル伝達機構において、EGFRを活性化するリガンドとして、ヘパリン結合性EGFファミリーに属するheparin-binding EGF-like growth factor(HB-EGF)に着目し、ヒト悪性グリオーマ細胞及び、悪性グリオーマ組織におけるHB-EGFの発現を検討し、その機能解析を行ったものであり、下記の結果を得ている。 1.HB-EGFcDNAをプローブとしたNorther blot解析にて、検討した7種類全てのヒト悪性グリオーマ細胞株で、HB-EGFmRNAの発現が認められた。また、ヒト悪性グリオーマ(glioblastoma)組織では、非腫瘍部脳に比較し2-5倍高いHB-EGFmRNAの発現を認めた。 2.グリオーマ細胞の細胞膜タンパクをビオチン化後、膜結合型proHB-EGFを特異的に認識する抗体を用いて免疫沈降し、HB-EGFタンパクの発現をWestem blot法を用いて解析したところ、約30kDaの膜結合型proHB-EGFに相当するバンドが認められた。以上より、グリオーマ細胞は、細胞表面に膜結合型proHB-EGFタンパクを発現していることが確認された。 3.悪性グリオーマ組織でのproHB-EGFの発現細胞を確認するために、そのレセプターとして機能することが知られているEGFRの発現とを併せて、それぞれの特異的抗体を用いた免疫組織化学染色法で検討した。抗EGFR抗体では、腫瘍細胞が染色され、血管内皮細胞は染色されなかった。抗proHB-EGF抗体では腫瘍細胞と、血管増生の部位での血管内皮細胞、及びpericyteと思われる細胞が陽性像を示した。検討した34例の悪性グリオーマ(anaplastic astrocytoma9例、glioblastoma25例)組織中、EGFRが陽性であったanaplastic astrocytomaの3例中2例(66%)で、また、glioblastomaの13例では全例(100%)でproHB-EGFも陽性であった。さらに、EGFR陰性のanaplastic astrocytomaの2例、glioblastomaの4例においても、proHB-EGFは陽性であった。 4.proHB-EGFは、ジャクスタクライン増殖因子として働くだけでなく、ジフテリア毒素(DT)のレセプターとしても機能していることが示されている。そこで、グリオーマ細胞で発現が確認されたproHB-EGFがDTのレセプターとして機能するかを、DT投与後のタンパク合成阻害能を指標に検討した。10ng/mlのDT投与によりproHB-EGFを発現しているグリオーマ細胞のタンパク合成能は、DT未投与のコントロール細胞の約20%まで阻害された。また、proHB-EGFの発現がないmouse L cellでは、DTによるタンパク合成阻害能は認められなかった。以上の結果は、proHB-EGFがDTレセプターとしても機能しうるというこれまでの報告に矛盾しないものであった。 5.グリオーマ細胞に対し、HB-EGFは増殖活性を有し、またオートクライン増殖因子として機能しうるかを検討した。血清除去下で培養したグリオーマ細胞に、種々の濃度でrecombinant HB-EGF(rHB-EGF)を投与した結果、濃度依存性に細胞増殖増強効果が認められ、10ng/mlのrHB-EGF投与では、未投与のコントロール細胞に比較し、細胞数が約2倍に増加することがわかった。一方、HB-EGFに対する特異的な中和抗体を加えると、コントロールのIgGを加えたものに比較し、約30-40%の細胞増殖抑制が認められた。以上の結果より、HB-EGFはグリオーマ細胞に対して増殖活性を有し、オートクライン増殖因子として機能していることが明らかとなった。 basic FGF(bFGF)及びEGFは、グリオーマ細胞に対して増殖因子として働くことが示されている。そこで、これらのグリオーマ細胞に対する増殖活性をDNA合成能を指標にHB-EGFのそれと比較した。10ng/mlのrHB-EGFの投与は、5%血清投与に比べると、1.2-2.3倍程度、グリオーマ細胞でのDNA合成能を促進することがわかった。rHB-EGFのもつDNA合成促進能は、10ng/mlのEGF,bFGFに比較すると、10-20%とわずかながら高かった。 6.グリオーマ細胞におけるHB-EGFの発現が、HB-EGF自体、あるいは他の増殖因子による発現制御を受けるかを、T98G細胞を用いて検討した。rHB-EGF投与により、またEGF,TGF 7.血管内皮特異的な増殖因子であるvascular endothelial growth factor(VEGF)は、グリオーマの血管新生に重要な役割を果たしていることが示されている。HB-EGFは、直接内皮細胞に対して増殖活性をもたないが、VEGFを誘導できれば、間接的に血管新生に関与すると考えられる。そこで、rHB-EGFをグリオーマ細胞に投与し、VEGFが誘導されるかを検討した。10ng/mlのrHB-EGFを投与すると、検討した3種のグリオーマ細胞株でVEGFmRNAの発現が亢進することがわかった。また、このVEGFmRNAの発現誘導は、加えるrHB-EGF濃度依存性に増加した。さらにrHB-EGFによるVEGFmRNAの発現誘導は、培養液中へのVEGFタンパク分泌促進につながることが、VEGF抗体を用いたWestem blot解析の結果より確認された。 以上、本論文はHB-EGF及びproHB-EGFがヒト悪性グリオーマ細胞、腫瘍組織で発現しており、TGF | |
| UTokyo Repositoryリンク | http://hdl.handle.net/2261/54038 |
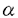 (TGF
(TGF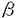 によってはこのような発現誘導は見られなかかった。
によってはこのような発現誘導は見られなかかった。