脳神経細胞の障害に起因する脳神経疾患においては、慢性的な経過、すなわち脳血管障害後の後遺症や神経変性疾患の進行に対する有効な治療法が少ない。私は、細胞内セカンドメッセンジャーであるcAMPが、ドパミン産生神経細胞に対する神経栄養効果などの多様な神経細胞機能を調節することに注目し、脳神経疾患の治療手段として細胞内におけるcAMP分解抑制が有望であると考えた。そこで、cAMP特異的分解酵素であるホスホジエステラーゼ(phosphodiesterase,PDE)4型選択的阻害薬ロリプラムが中枢神経細胞に及ぼす効果について、特にドパミン神経細胞に注目して検討し、脳神経疾患治療薬としてのロリプラムの利用意義について考察した。 ホ乳類の脳にはPDE1、PDE2、PDE3およびPDE4が存在すると報告されている。はじめに、神経細胞内cAMP量調節におけるPDE4の存在意義を他のタイプのPDEと比較するため、初代培養したラット大脳皮質神経細胞中のcAMPをエンザイムイムノアッセイ(EIA)によって定量することにより、アドレナリン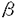 受容体刺激条件下における細胞内cAMPレベルに対するロリプラム及びその他の各種PDE阻害薬の効果について検討した。PDE4阻害薬であるロリプラムとRo20-1724、およびPDE2阻害薬であるEHNAは、非刺激条件下においてはcAMP量に影響を及ぼさなかったが、イソプロテレノール誘発性のcAMP上昇を有意に増幅した。ビンポセチン(PDE1阻害薬)、ミルリノン(PDE3阻害薬)およびザプリナスト(PDE5及びPDE6阻害薬)は非刺激条件下および刺激条件下のいずれにおいても、細胞内cAMP量に影響を及ぼさなかった。 受容体刺激条件下における細胞内cAMPレベルに対するロリプラム及びその他の各種PDE阻害薬の効果について検討した。PDE4阻害薬であるロリプラムとRo20-1724、およびPDE2阻害薬であるEHNAは、非刺激条件下においてはcAMP量に影響を及ぼさなかったが、イソプロテレノール誘発性のcAMP上昇を有意に増幅した。ビンポセチン(PDE1阻害薬)、ミルリノン(PDE3阻害薬)およびザプリナスト(PDE5及びPDE6阻害薬)は非刺激条件下および刺激条件下のいずれにおいても、細胞内cAMP量に影響を及ぼさなかった。 これらのことより、初代培養した大脳皮質神経細胞において、アドレナリン受容体刺激によって増加するcAMPを分解する主要なPDEはPDE4およびPDE2であることを見出した。また、神経細胞内cGMPレベルに対しては、EHNAのみが細胞内cGMPレベル上昇をもたらし、ロリプラムは無影響であった。このことより、ロリプラムが神経細胞内のcAMP量を選択的に上昇させる上で効果的な薬物であることを確認した。大脳皮質神経細胞、アストロサイト及び血管平滑筋細胞の細胞内cAMPレベルについて検討したところ、ロリプラムは、いずれの細胞においてもフォルスコリン存在下で細胞内cAMPレベルを上昇させたが、最低有効濃度は細胞ごとに異なった(図-1)。これらの知見は、細胞の種類によるPDE4サブタイプの多様性を示唆している。すなわち、中枢神経細胞にはロリプラムに対する感受性の高いPDE4が存在し、cAMP代謝調節機構において重要な役割を果たしていると考えられた。 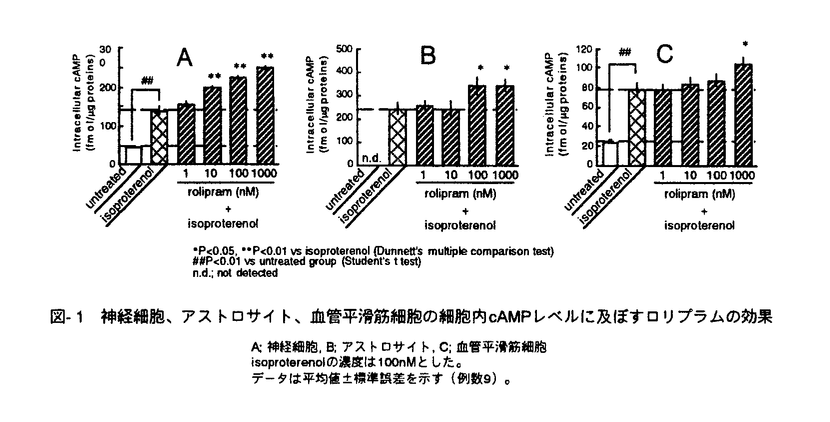 図-1 神経細胞、アストロサイト、血管平滑筋細胞の細胞内cAMPレベルに及ぼすロリプラムの効果A;神経細胞,B;アストロサイト,C;血管平滑筋細胞isoproterenolの濃度は100nMとした。データは平均値±標準誤差を示す(例数9)。 図-1 神経細胞、アストロサイト、血管平滑筋細胞の細胞内cAMPレベルに及ぼすロリプラムの効果A;神経細胞,B;アストロサイト,C;血管平滑筋細胞isoproterenolの濃度は100nMとした。データは平均値±標準誤差を示す(例数9)。 次に、初代培養したラット中脳神経細胞を用いて、ドパミン及びその代謝物(ジヒドロキシフェニル酢酸,DOPAC)の細胞内含量あるいは細胞外放出量を高速液体クロマトグラフィー(HPLC)により測定し、ドパミン生合成に対するロリプラムの作用を、他のタイプのPDEに対する選択的阻害薬と比較した。ロリプラムはフォルスコリン非存在下では細胞内ドパミン及びDOPACレベルに影響を及ぼさなかったが、フォルスコリンの誘発する細胞内ドパミン及びDOPACレベルの増加を有意に増強した(図-2)。これに対し、ビンポセチン、EHNA、ミルリノン、ザプリナストは、フォルスコリンによる刺激の有無に関わらず、ドパミンおよびDOPACレベルを増加させなかった。細胞外のドパミンおよびDOPACレベルに及ぼす効果については、ロリプラムとRo20-1724が、フォルスコリン刺激条件下において細胞外ドパミンおよびDOPACレベルを有意に増加させた。このことは、ロリプラムによるドパミン含量増加が、単に放出抑制によって貯溜したものではなく、ドパミン生合成を高めた結果であることを示している。 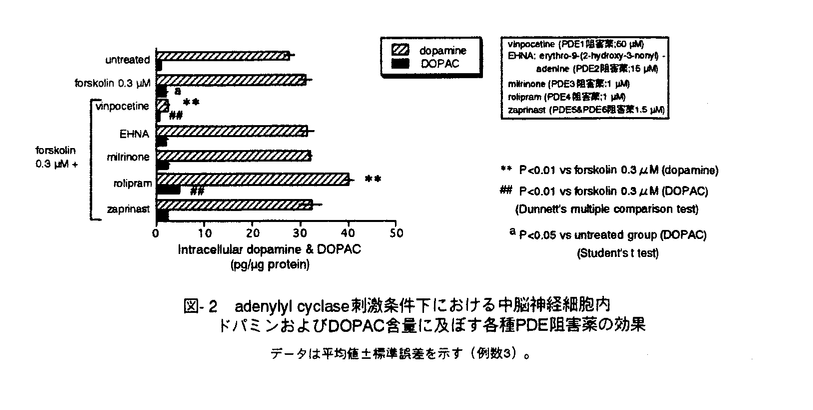 図-2 adenylyl cyclase刺激条件下における中脳神経細胞内ドパミンおよびDOPAC含量に及ぼす各種PDE阻害薬の効果データは平均値±標準誤差を示す(例数3)。 図-2 adenylyl cyclase刺激条件下における中脳神経細胞内ドパミンおよびDOPAC含量に及ぼす各種PDE阻害薬の効果データは平均値±標準誤差を示す(例数3)。 次に、ラット中脳ドパミン産生細胞の生存に及ぼすロリプラムの影響を、チロシンヒドロキシラーゼ(ドパミン生合成経路の律速酵素)発現細胞数を指標として検討し、微小管関連タンパク質タイプ2(MAP2)、グルタミン酸、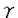 -アミノ酪酸(GABA)のそれぞれを発現する細胞の数に及ぼす効果と比較した。フォルスコリン存在下、ロリプラムは10nM以上の濃度において、TH陽性細胞の生存を促進させた(図-3)。また、ロリプラムは10 -アミノ酪酸(GABA)のそれぞれを発現する細胞の数に及ぼす効果と比較した。フォルスコリン存在下、ロリプラムは10nM以上の濃度において、TH陽性細胞の生存を促進させた(図-3)。また、ロリプラムは10 Mの濃度においてMAP2陽性細胞数も増加させたが、グルタミン酸陽性細胞数及びGABA陽性細胞数には影響を及ぼさず、ロリプラムによって生存が増強される神経細胞は主としてドパミン産生神経細胞であると考えられた。 Mの濃度においてMAP2陽性細胞数も増加させたが、グルタミン酸陽性細胞数及びGABA陽性細胞数には影響を及ぼさず、ロリプラムによって生存が増強される神経細胞は主としてドパミン産生神経細胞であると考えられた。 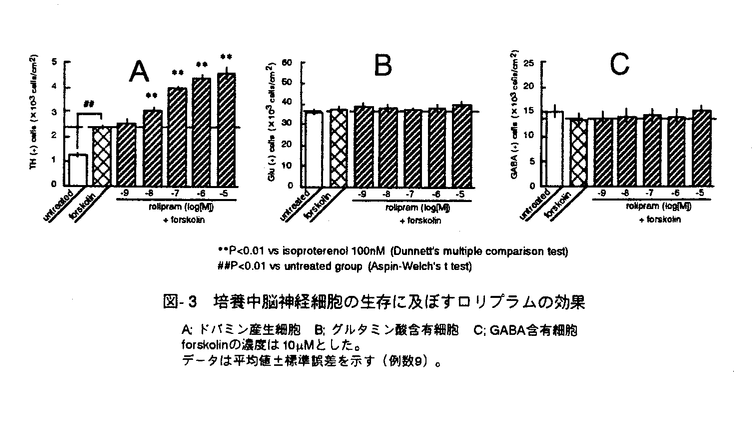 図-3 培養中脳神経細胞の生存に及ぼすロリプラムの効果A;ドパミン産生細胞 B;グルタミン酸含有細胞 C;GABA含有細胞forskolinの濃度は10Mとした。データは平均値±標準誤差を示す(例数9)。 図-3 培養中脳神経細胞の生存に及ぼすロリプラムの効果A;ドパミン産生細胞 B;グルタミン酸含有細胞 C;GABA含有細胞forskolinの濃度は10Mとした。データは平均値±標準誤差を示す(例数9)。 最後に、ロリプラムの効果をin vivoにおいて確認するため、ラット脳内カテコールアミン系神経伝達物質量に対する効果を検討した。ラットにロリプラム0.3mg/kgまたは溶媒を1日1回、14日間反復経口投与した後、マイクロウェーブを照射した脳組織中のモノアミン含量を、HPLCを用いて測定した。単回経口投与においては、ロリプラムは検討したいずれの部位のドパミン量についても有意な変化を示さなかったが、反復経口投与によって前頭皮質のドパミン含量を対照群(精製水投与)の2.4倍にまで有意に増加させた(図-4)。他の部位のドパミン量は、反復投与においても、対照群と比較して有意な差は認められなかった。またノルアドレナリン量は、単回投与、反復投与のいずれの場合も検討した全ての部位において有意な変化が認められなかった。以上の結果より、ロリプラムはin vivoにおいてもドパミン量を増加させることが示された。また、ドパミン含量増加に部位差が確認されたことより、ロリプラムは特定のドパミン神経系に効果を発揮する可能性が考えられた。 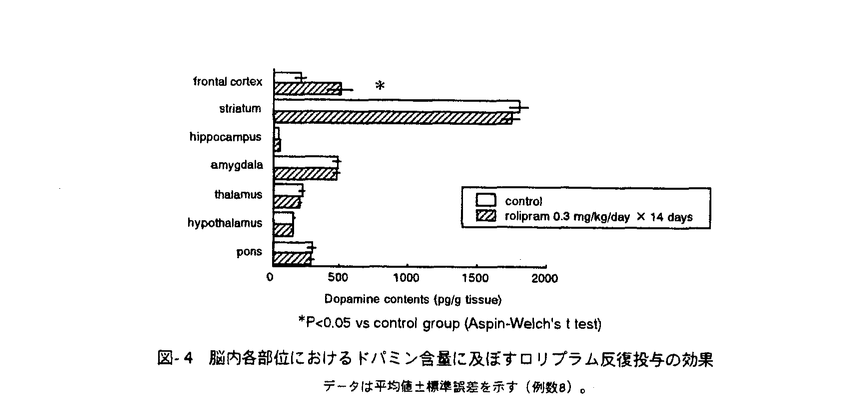 図-4 脳内各部位におけるドパミン含量に及ぼすロリプラム反復投与の効果データは平均値±標準誤差を示す(例数8)。 図-4 脳内各部位におけるドパミン含量に及ぼすロリプラム反復投与の効果データは平均値±標準誤差を示す(例数8)。 私は本研究において、ロリプラムが10nMという低濃度でPDE4阻害による中枢神経細胞のcAMPレベル上昇をもたらすこと、及びロリプラムがドパミン産生神経細胞に対し、同様の低濃度においてドパミン生合成促進及び生存促進をもたらすことを見出した。さらに、ロリプラム反復投与によりラット脳内ドパミンが部位選択的に増加することを示し、ロリプラムがin vivoにおいても、ドパミン産生細胞に対する賦活効果を発揮することを確認した。ロリプラムは神経細胞選択的に、かつ伝達物質や生存など様々な神経機能を賦活させるユニークな脳神経疾患治療薬になりうる。特に出血部位が安定していない脳血管障害後遺症患者やドパミン神経系に障害が認められる疾患の治療への応用が期待される。また、ロリプラムの作用を追究する過程において、神経細胞の機能及び生存に及ぼすPDE1、PDE2及びPDE4阻害薬がそれそれ特徴的な作用を示したことから、脳におけるそれぞれのPDEの役割分担のパターンを見ることができた。これらの知見は、今後の中枢神経細胞の各種機能における環状ヌクレオチドの役割についての研究において、有用な情報と考えられる。 |