本研究は神経細胞における細胞内物質輸送のメカニズムを明らかにするため、キネシン型モーター分子のコンセンサスシークエンスを利用して新たな分子(KIFC2)を発見、その機能解析を試みたものであり、以下の結果を得ている。 1.キネシンスーパーファミリー蛋白質(KIF)のアミノ酸一次構造を比較することにより、C末モータードメイン型KIFCに特異的なコンセンサスシークエンスを発見した。このコンセンサスシークエンス利用して、二つの新たなKIFCであるKIFC1とKIFC2をクローニングした。KIFC2は系統発生学的解析により全く新しいKIFであることが示された。 2.他のKIFとの一次構造の比較、抗KIFC2抗体によるnative KIFC2の免疫精製で関連蛋白質は検出されなかったこと、ショ糖密度勾配遠心法での他のKIFとの沈降係数の比較により、KIFC2の分子構造はホモ二量体と推測されることが示された。 3.KIFC2のモーター活性を証明するin vitro motility assayには、大腸菌とバキュロウイルスの発現系でrecombinant KIFC2が不溶性の細胞内封入体を形成するために成功していない。しかしながらKIFC2のdeletion mutantが微小管依存性ATPase活性を持つこと、生理的なイオン強度でヌクレオチド依存性に微小管と結合、解離能を有することを示し、KIFC2が微小管依存性のモーターであることを示唆した。 4.mRNAブロッティングと免疫ブロッティングでKIFC2の発現を解析したところ、KIFC2は神経系に特異的に発現し、胚ではほとんど検出されず、成長に伴い増加し、成獣脳に豊富に発現していることを示した。この発現パターンは有糸分裂のモーターとは全く異なり、KIFC2が細胞分裂に関わるのではないことを示した。 5.抗KIFC2抗体を作成し、マウス脳の免疫ブロッティング、およびマウス脳の免疫染色を行ったところ、KIFC2が神経細胞の特に細胞体や樹状突起に局在し、軸索には存在しないことを示した。さらに遺伝子組換えアデノウイルスベクターによりmyc-tagged KIFC2を培養神経細胞で過剰発現したところ、KIFC2がやはり樹状突起と細胞体に局在し、軸索には検出されないことを示し、KIFC2が樹状突起の輸送モーターであることを明らかにした。 6.培養海馬神経細胞の蛍光染色で、抗KIFC抗体で染まる大型班点状構造が、細胞内オルガネラのマーカーであるミトコンドリアのMitotracker、シナプス小胞やその前駆体のsynaptophysin、RNA顆粒のSYTO14と共染しないことを示し、KIFC2がこれらのオルガネラを輸送するのではないことを示した。 7.大脳皮質抽出液を分画遠心法、及びNycodenz密度勾配遠心法(NDG)で分画後、免疫沈降法でKIFC2を含む膜性小器官を精製し、KIFC2cargoの分子構成を解析したところ、KIFC2cargoは他の微小管依存性モーター(KIF1A、KIF3、KIF5、cytoplasmic dynein、dynactin)、シナプス小胞蛋白質(synaptophysin,SV2)、シナプス前膜蛋白質(syntaxin,SNAP-25)、シナプス後膜蛋白質(glutamate receptor)、被膜小胞に関係する蛋白質(clathrin,adaptinb,adapting)、エンドサイトーシスに関係する蛋白質(dynamin,rab5,Lamp2,trk)等と共沈しないことを示し、これらの蛋白を含むオルガネラを輸送するのではないことを示した。 8.KIFC2を結合した膜性小器官を抗KIFC2抗体で免疫精製し、電子顕微鏡で観察したところ、ほとんどが多胞体様のオルガネラであることを示し、これを輸送していることを明らかにした。このオルガネラは、Rhodamine-BSAラベル、transferrin receptor、Lamp2を含まず、既知のエンドソームとは異なる新しいクラスの多胞体であることを示した。 以上のように本論文は細胞内物質輸送に関わる初のC末モータードメイン型KIFを発見し、神経細胞樹状突起における多胞体様の細胞内小器官を輸送するモーターであることを明らかにした。本研究は神経細胞における細胞内物質輸送のメカニズムの解明に重要な貢献をなすと考えられ、学位の授与に値するものと考えられる。 | 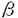 1strandのすぐ上流の頸部に発見した。このコンセンサスシークエンス利用して、二つの新たなKIFCであるKIFC1とKIFC2をクローニングした。KIFC1はCHO2antigenのホモログであった。
1strandのすぐ上流の頸部に発見した。このコンセンサスシークエンス利用して、二つの新たなKIFCであるKIFC1とKIFC2をクローニングした。KIFC1はCHO2antigenのホモログであった。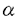 -helical coiled coil構造をとり、N末端側は尾部を、C末端側はモータードメインを形成すると予測され、kar3/ncdサブファミリーと同様にホモダイマーではないかと予測された。また、ショ糖密度勾配遠心法の結果、nativeのKIFC2蛋白の沈降係数は6Sであり、
-helical coiled coil構造をとり、N末端側は尾部を、C末端側はモータードメインを形成すると予測され、kar3/ncdサブファミリーと同様にホモダイマーではないかと予測された。また、ショ糖密度勾配遠心法の結果、nativeのKIFC2蛋白の沈降係数は6Sであり、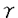 )、エンドサイトーシスに関係する蛋白質(dynamin,rab5,Lamp2,trk)などの多くの神経細胞蛋白質をテストしたがKIFC2と共沈しなかった。
)、エンドサイトーシスに関係する蛋白質(dynamin,rab5,Lamp2,trk)などの多くの神経細胞蛋白質をテストしたがKIFC2と共沈しなかった。