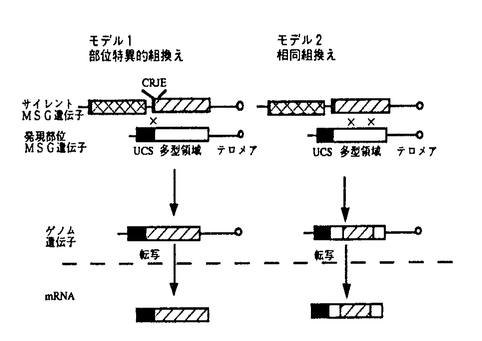
本研究はAIDSなどの合併症として重要なカリニ肺炎の病原体ニューモシスチス・カリニの主要表面抗原(MSG)について分子生物学的方法で解析を行ったものであり、下記の結果を得ている。 1.イムノスクリーニングによってMSGcDNAをクローニングし塩基配列を解析したところ、MSGcDNAは多型なサブタイプで構成されるファミリーであることが明らかとなった。サブタイプ間の相同性(identity)はアミノ酸レベルで約70%であり、約4kbの長さのMSGcDNAのほとんどの領域に多型性が認められる。しかしながら、最も5’領域の約400bpの配列(UCS)のみは保存されていることが判明し、UCSに多型領域が続くというMSGcDNAの一般構造が示された。 2.MSGゲノム遺伝子についてパルスフィールド電気泳動による染色体DNAの解析、サザンブロット解析などを行ったところ、MSGゲノム遺伝子は多型多重であるが、cDNAに保存されたUCSを持つMSG遺伝子は、約14本の染色体の内、約500kbの長さの1本の染色体上にのみ存在していた。このUCSを持つMSG発現部位遺伝子はエキソヌクレアーゼによる消化実験及びクローニングした遺伝子の解析からテロメア近傍に位置していることが示された。その他の多数のMSG遺伝子はUCSを持たず非発現遺伝子と考えられるが、非発現遺伝子はその多くがゲノム上でタンデムに並ぶクラスターを形成しており、少なくともその一部がテロメア近傍に位置していた。 UCSを持つ発現部位遺伝子を複数クローニングして解析したところ、得られたクローンはUCSの下流に各々異なる多型領域が直結していたことから、UCS部位におけるDNA組換えによりcDNA多型が形成されているものと考えられた。 3.分類学的にニューモシスチス・カリニに近縁で発現解析の可能な分裂酵母において、UCS上流領域がプロモーターとして機能し得るかを検討したところ、UCS上流領域にプロモーター活性が認められ、UCS部位がMSGの発現部位であることが示唆された。 4.多数のMSGcDNA及びMSG遺伝子の塩基配列の比較を行い、多型性発現にはたらくメカニズムを検討したところ、発現部位であるUCSを持つMSG遺伝子と非発現部位のMSG遺伝子の間には28bpの配列(CRJE)が保存されており、CRJEでの部位特異的組換えが示唆された。また、多型領域における比較において完全に一致する領域と類似領域を合わせ持つキメラな遺伝子が認められ、多型領域における相同組換えが示唆された。これらのCRJEにおける部位特異的組換えと多型領域における相同組換えが協調してはたらき、発現部位のMSG遺伝子が変換されて異なるMSG遺伝子が発現することにより、MSGが多型となっているというモデルが提唱された。 以上、本論文はニューモシスチス・カリニのMSGの分子生物学的解析によって、MSGの遺伝子レベルでの多型性を発見し、MSGの多型性が特異的なDNA組換えによって遺伝的に制御されている可能性が高いことを示した。表面抗原分子の多型性とその遺伝的制御機構の存在を明らかにしたことは、ニューモシスチス・カリニの病原性の解明に重要な貢献をなすものと考えられ、学位の授与に値するものと考えられる。 |