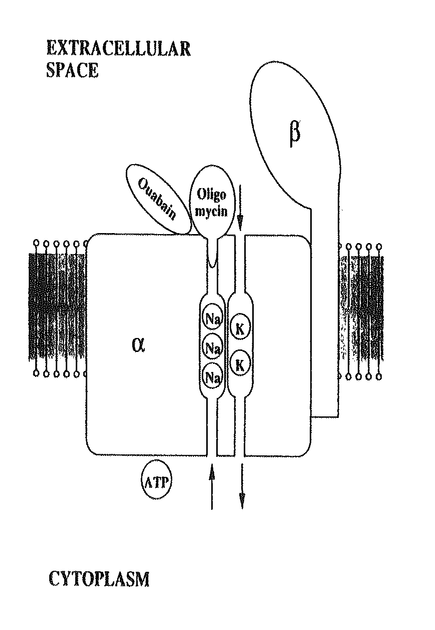
Na/K-ATPaseはNa-ポンプとも呼ばれ、動物細胞に普遍的に存在しNa+とK+の能動輸送を担っている膜蛋白質である。本研究は抗生物質のひとつオリゴマイシンのNa/K-ATPaseに対する阻害作用を検討することによって、Na/K-ATPaseのイオン輸送機構の解明を試みたものであり、下記の結果を得ている。 1.オリゴマイシンはNa/K-ATPase活性やイオン輸送を阻害し、Na+結合を促進するが、K+結合やK+輸送には影響をあたえなかった。そしてその阻害曲線ならびに促進曲線がすべて一相性であること、これらのI50ならびにK0.5が近似した値を示したことから、この薬剤の特異的結合部位は反応速度論的に一種類であることが示された。 2.オリゴマイシン存在下、非存在下でのNa/K-ATPaseへのNa+、K+結合を測定しその解離定数を比較したところ、オリゴマイシンはNa+の親和性を増大させるが、K+の親和性には影響を与えないことが示された。 3.種々のリガンド存在下におけるNa/K-ATPaseのトリプシン限定分解パターンを観察した結果、Na+、K+、ATP、ウワバインと異なり、オリゴマイシンはATPaseのコンホメーション変化をおこさないことが示された。 4.オリゴマイシンはNa/K-ATPaseの細胞質側に結合部位があるATPの結合にほとんど影響をあたえなかったが、細胞外側に結合部位があるウワバインの結合を阻害した。しかし、それは競合阻害ではないことが示された。また、ウワバイン低感受性ならびに高感受性Na/K-ATPaseアイソフォームを、オリゴマシンは同様に阻害した。したがって、この薬剤の結合部位は細胞外側であるがウワバインの結合部位とは異なることが示された。 5.以上の結果から、「オリゴマイシンはNa/K-ATPaseのコンホメーション変化を介するのではなく、ATPaseの細胞外側からNa+の遊離部位またはその近傍に結合し、Na+の遊離を直接阻害することによって見かけ上Na+の結合量を増大させNa+輸送およびATPase活性を阻害する」との結論が導かれ、これをふまえてNa+とK+の輸送路が異なっているというモデルが提唱された。 以上、本論文はNa/K-ATPaseに対するオリゴマイシンの阻害機構の詳細な研究に基づいて、Na+、K+の輸送路が異なるという新しい概念を提唱している。イオンと膜蛋白質の相互作用は、細胞内への有機物質の取り込み、さらには細胞内および細胞間の情報伝達と深くかかわっているが、その相互作用の仕組みについては不明な点が多い。本論文はNa/K-ATPaseを用いてこの課題に取り組み、新しいイオン輸送モデルの提唱に到達しており、その結果はこの分野の研究に重要な貢献をして、学位の授与に値するものと考えられる。 |  鎖と55kDaの
鎖と55kDaの 鎖の二つのサブユニットから成る膜タンパク質で、
鎖の二つのサブユニットから成る膜タンパク質で、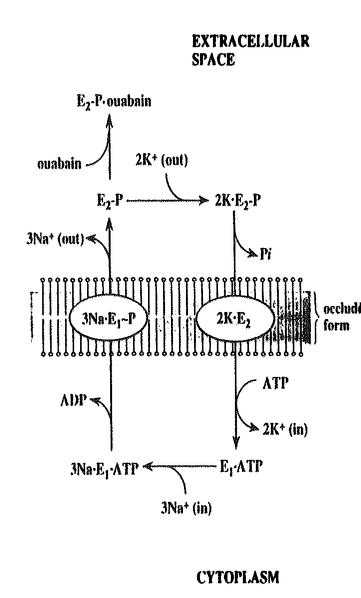
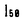 )は2.2
)は2.2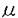 Mであった。また、この薬剤はNa-KおよびNa-Na交換輸送反応を各々
Mであった。また、この薬剤はNa-KおよびNa-Na交換輸送反応を各々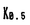 )は4.5
)は4.5