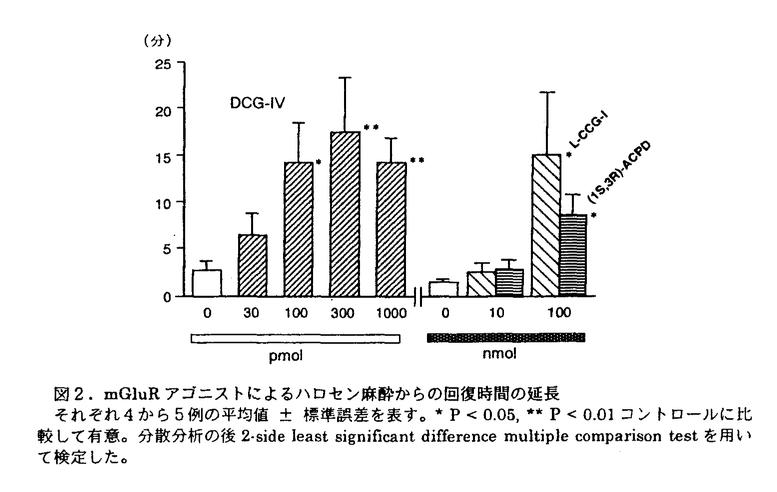
L-グルタミン酸は哺乳類中枢神経系の主な興奮性神経伝達物質の一つと考えられており、その受容体には、イオンチャンネル型グルタミン酸受容体(iGluR)と、代謝調節型グルタミン酸受容体(mGluR)の2種類があることが知られている。様々なアゴニスト・アンタゴニストが開発されている事などから、iGluRは比較的研究が進んでいる。mGluRは現在までに8つのサブタイプ(mGluR1〜mGluR8)がクローニングされており、2次メッセンジャー系の差・アゴニストの選択性の違いにより、GroupI(mGluR1,5)・Group II(mGluR2,3)・Group III(mGluR4,6,7,8)の3つのグループに細分されている。選択的アゴニスト・アンタゴニストの開発が遅れたこともあり、mGluRの機能には不明な点が多かった。近年篠崎らは、2-(carboxycyclopropyl)glycine(CCG)の誘導体から数種の強力なmGluRアゴニストを開発した。その中の一つ、(2S,1’R,2’R,3’R)-2-(2,3-dicarboxycyclopropyl)glycine(DCG-IV)はGroup II mGluRの選択的且つ強力なアゴニストであり、膜電位感受性Ca2+電流を抑制して神経終末からの神経伝達物資の遊離を抑制することが、in vitroの実験から明らかにされている。本研究は、これらのアゴニストを用いて、特にin vivoにおける実験を中心に、mGluRの生理機能の解明を行ったものである。 1.ラットにおけるmGluRアゴニストによるハロセン麻酔の睡眠延長 まず、mGluRアゴニストを投与した際のラットの行動変化を調べた。DCG-IVを側脳室内に投与すると、極く低用量の30-300pmolで用量依存的にハロセン麻酔からの回復時間を延長させた。用量を1nmolにすると弱い中枢興奮作用が発現し、ハロセン麻酔からの回復時間延長作用は逆に減弱した。即ち、DCG-IVの中枢抑制作用はベル型の用量作用曲線を示した。非選択的mGluRアゴニスト(1S,3R)-1-amino-cyclopentane-1,3-dicarboxylic acid[(1S,3R)-ACPD]や、Group II mGluRアゴニストとしての効力や選択性がDCG-IVより低い(2S,1’S,2’S)-CCG(L-CCG-I)は、100nmolではじめて回復時間延長作用が認められた。この中枢抑制作用の強さはin vitroで報告されているGroup II mGluRアゴニスト活性と平行していた。即ち、Group II mGluRを活性化すると中枢抑制作用を示すことが明らかとなった。 2.DCG-IVにより誘発される中枢興奮と神経細胞死 DCG-IVの投与量を上げた際の中枢興奮作用について検討した。DCG-IVの投与量を50nmolに上げると、痙攣及び部位特異的な神経細胞死が誘発された。この作用はNMDA受容体の活性化を介したものとも考えられた。しかし、NMDAの興奮毒性を抑制できる量の、NMDA受容体アンタゴニストである3-[(RS)-2-carboxy-piperazin-4-yl]propyl-1-phosphonic acid(CPP)では、DCG-IV50nmolの中枢興奮作用は一部しか抑制されず、完全に抑制するには高用量のCPPを要した。従ってDCG-IVの中枢興奮作用はNMDA受容体を活性化するのみでなく、Group II mGluRとNMDA受容体の活性化の協調によって誘発されている可能性が考えられた。 3.カイニン酸の興奮毒性に対するDCG-IVの抗痙攣及び神経細胞保護作用 低用量のDCG-IVによる中枢抑制作用を更に検討するため、カイニン酸(KA)を投与した際に見られる痙攣や神経細胞死に対する作用について検討した。KAをラットの脳室内に投与すると、wet-dog shakes(WDS)などの興奮状態がみられ、1時間程すると辺縁系痙攣が数分間隔で起こり(散発的辺縁系痙攣)、時間と共にこの辺縁系痙攣が絶え間なく起こるようになった(持続的辺縁系痙攣)。KA投与1週間後には、海馬CA3・大脳辺縁系・膝状体内側部で神経細胞死が観察された。DCG-IVの単回投与では、KAによる痙攣を抑制することができなかったため、浸透圧ポンプを用いて、低用量のDCG-IVを持続的に側脳室内に適用しながら、KAを投与した。DCG-IVの持続投与それ自体では、ラットに目立った行動上の変化は観察されなかった。KA投与によるWDSや散発的辺縁系痙攣の発現は抑制されなかったが、DCG-IV80pmol/hにより持続的辺縁系痙攣の発現が有意に抑制された。また、DCG-IV24,80pmol/hの持続投与により大脳辺縁系・膝状体内側部でのKA誘発神経細胞死が軽減された。更に、KAに最も脆弱である海馬CA3領域の錐体細胞の変性も軽減された。即ち、DCG-IVによるGroup II mGluRの持続的な活性化は、KAによる痙攣を減弱させ、部位特異的神経細胞保護作用を示すことが明らかとなった。 4.ラット扁桃核キンドリングに対するDCG-IVの抗痙攣作用 Group II mGluRの活性化が抗痙攣作用を示したため、ラット扁桃核キンドリングに対するmGluRアゴニストの抗痙攣作用を検討した。L-CCG-I(500nmol)の脳室内投与ではキンドリングの発達は抑制されなかったが、完成したキンドリング痙攣がL-CCG-I投与により完全に抑制される例が見られた。DCG-IV80pmol/hを持続的に投与しながら扁桃核刺激を行ってもキンドリングは正常に発達した。一方、キンドリングが完成したラットにDCG-IV80pmol/hを持続投与すると、扁桃核刺激による痙攣が軽減された。以上から、GroupIImGluRの活性化はキンドリング痙攣の発達には影響せず、完成したキンドリング痙攣を抑制することが示唆された。 以上よりGroup II mGluRアゴニストは中枢抑制作用を示すことが明らかとなった。DCG-IVは極めて低用量で中枢抑制作用を示したが、意識の喪失を伴うような非特異的なものではなく、それ自体では一般行動に大きな影響を与えないものであった。更に、DCG-IVは一般行動に影響を与えない程度の低用量の持続投与により、部位特異的な神経細胞保護作用や抗痙攣作用を示した。抗痙攣作用も完成したキンドリング痙攣やKA誘発の持続的辺縁系痙攣のような特定の場合にのみ顕著に現れる可能性が示された。これらの作用は、Group II mGluRの活性化により、神経終末からの神経伝達物質(グルタミン酸)の過剰な遊離を抑制するためと考えられる。 本研究の成果は、代謝調節型グルタミン酸受容体の生理学的機能の解明に大いに貢献し、Group II mGluR活性化の観点からの新たな抗痙攣薬・神経保護薬などの創薬に寄与すると考えられ、博士(薬学)の学位を授与するに値すると認めた。 | 
 molの高用量を投与すると、軽度の中枢興奮作用が観察されたものの、神経細胞死は見られなかった。高い用量のDCG-IVはNMDA受容体も活性化することが明らかにされており、高用量で観察された中枢興奮作用が、NMDA受容体の活性化を介したものであることも考えられた。そこでNMDAの興奮毒性を抑制できる十分な量(300pmol)の、NMDA受容体の競合的アンタゴニストである3-[(RS)-2-carboxypiperazin-4-yl]propyl-1-phosphonic acid(CPP)を同時に投与しても、DCG-IV50nmolの中枢興奮作用は一部しか抑制する事ができず、完全に抑制するためにはCPP3nmol以上必要であった。従ってDCG-IVの中枢興奮作用は単にNMDA受容体を活性化するのみでなく、Group II mGluRとNMDA受容体の活性化の協調によって誘発されている可能性が考えられた。
molの高用量を投与すると、軽度の中枢興奮作用が観察されたものの、神経細胞死は見られなかった。高い用量のDCG-IVはNMDA受容体も活性化することが明らかにされており、高用量で観察された中枢興奮作用が、NMDA受容体の活性化を介したものであることも考えられた。そこでNMDAの興奮毒性を抑制できる十分な量(300pmol)の、NMDA受容体の競合的アンタゴニストである3-[(RS)-2-carboxypiperazin-4-yl]propyl-1-phosphonic acid(CPP)を同時に投与しても、DCG-IV50nmolの中枢興奮作用は一部しか抑制する事ができず、完全に抑制するためにはCPP3nmol以上必要であった。従ってDCG-IVの中枢興奮作用は単にNMDA受容体を活性化するのみでなく、Group II mGluRとNMDA受容体の活性化の協調によって誘発されている可能性が考えられた。