| 内容要旨 | | 緒言 哺乳動物においては,胎盤は母体から胎児へ酸素および栄養の供給を行ない,胎児を発育させるという重要な機能を有している.一方,胎盤を構成している主な細胞であるtrophoblastは多くのホルモンを産生していて,生殖生理学において重要な位置をしめている. ヒト胎盤を構成する主な細胞である絨毛細胞(trophoblast)はhCG(human chorionic gonadotropin),hPL(human placental lactogen),GnRH(gonadotropin releasing hormone),CRF(chorticotropin releasing factor),ACTH,variant growth hormone(variant GH),PTH関連蛋白(PTHrP)などのペプチドホルモン,およびエストロジェン,プロゲステロンなどのステロイドホルモンを産生している.これらのホルモンは母体,胎盤自身あるいは胎児に作用し,妊娠の維持,胎児への経胎盤的カルシウム輸送,あるいは胎児の発育などに関与していると考えられている.一種類の細胞でこのように多種のホルモンを産生するのは絨毛細胞のみである.これらホルモンの産生調節機序および生理作用に関しては多くの研究がなされている.しかしながら,いまだ充分に解明されたとはいえない. hCGは胎盤ホルモンの代表的なものであり,その産生,分泌調節に関して多くの研究が蓄積されている.すなわち,絨毛癌細胞株,正常絨毛細胞の初代培養による実験系では,EGF,ハイドロコーチゾン,GnRH,cAMP,phorbor esterがhCGの産生分泌を促進すると報告されている.また,絨毛細胞が産生する主要なステロイドホルモンであるプロゲステロン(P)はcAMPにより分泌が促進されることが知られているが,first messengerはいまだ不明である. レチノール酸(retinoic acid-RA)はビタミンAの活性物質であり、元来催奇形物質として知られていたが,その後の研究により胚発生,胎児の神経系の分化,細胞分化およびさまざまな遺伝子の発現など,多種の細胞で多彩な作用を有していることが明らかにされつつある.また,RAの分化誘導作用を利用して前骨髄性白血病の治療薬として,現在臨床応用されている. 絨毛細胞におけるレチノール酸(RA)の作用に関する研究は少ない.わずかに,ある種の絨毛癌細胞株(JEG-3細胞)において,hCGおよびhCG-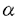 サブユニット(hCG-)の分泌を促進するという報告がある.一方,SV-40でトランスフォームした妊娠末期絨毛細胞においては,RAはhCGの分泌を促進するがhCG-の分泌を抑制すると報告されている.また,P分泌に関するRAの関与は言及はなされていない. サブユニット(hCG-)の分泌を促進するという報告がある.一方,SV-40でトランスフォームした妊娠末期絨毛細胞においては,RAはhCGの分泌を促進するがhCG-の分泌を抑制すると報告されている.また,P分泌に関するRAの関与は言及はなされていない. そこで,3種の代表的な絨毛癌細胞株(JEG-3,Jar,BeWo)を用いてRAのhCGおよびhCG-の分泌促進作用が絨毛癌細胞株に共通するものであるか,また,RAはP分泌になんらかの影響を及ぼすか,更に,正常絨毛細胞におけるRAのhCGあるいはP分泌促進作用の有無について検討した. 材料と方法1.絨毛癌細胞の培養 JEG-3細胞(ATCC)およびJar細胞(ATCC)は10%FCS、ペニシリン、ストレプトマイシン加DMEMで継代培養した.BeWo細胞(ATCC)は10%FCS、ペニシリン、ストレプトマイシン加Ham’sF10で継代培養した.0.5-1.0×105個の細胞を6穴multidishに撒き、1-2日後に培養液を除去し、新鮮な4%FCS加培養液を加えてRA,8-bromo-cAMPあるいはcholera toxinの添加実験を行なった.培養液の交換は24-48時間ごとに行なった.実験の最後に培養液を採取した.細胞は0.025%トリプシン-0.02%EDTAで剥がし細胞数を測定した.すべての実験はtriplicateで培養し,最低2回繰り返し行なった. 2.正常絨毛細胞の単離と培養 Klimanらの方法を用いて,妊娠初期および末期胎盤よりcytotrophoblastを得た.培養および添加実験は1.と同様な方法で行なった. 3.ホルモン測定および蛋白定量 a)培養液中のhCG,hCG-およびPの測定はRIAにより行なった. b)cAMPの測定 培養液を除去した後に,細胞内cAMPを65%エタノールで抽出した.抽出液を蒸発させRIA bufferで溶解した後RIAで測定した. c)蛋白定量 1M NaOHに溶解した0.2%triton-Xで細胞を可溶化した後、1M HClで中和、Bradfordの方法により蛋白定量を行なった. 4.細胞数の測定はCoulter社製Cell counterを用いて行なった.5.検定 すべての値はm±SEで表わした. 統計的有意差検定はanalysis of varianceあるいはStudent t-testにより行なった.p<0.01を有意差があると判定した. 成績1.JEG-3細胞におけるRAの影響 RA(10-6M)は3日間の添加でhCG,hCG-およびPの分泌を統計的有意差をもって促進した(表1).RAの分泌促進作用はRA添加後48時間目に現われ,添加3-4日目に最大の分泌促進作用を示した.P分泌促進作用はRA添加後3日目に出現し,5日目に最大の分泌促進作用を示した.4日間のRA添加では,RAは濃度依存的にこれらのホルモン分泌を促進し,10-8Mの濃度から統計的に有意差を有し,10-6Mで最大となった.cholera toxin(10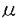 g/L)あるいは8-bromo-cAMP(1mM)は24時間の添加でhCG,hCG-およびPの分泌を統計的有意差をもって促進した.その分泌促進作用はRAより著明であった.RAとcholera toxinの両者の添加実験により,hCGおよびhCG-分泌は相乗的に促進された.また,P分泌は相加的に促進された.細胞内cAMP含量はRA添加により促進されなかった(表1).RA(10-10-5×10-6M)は細胞増殖あるいは細胞蛋白含量に影響を与えなかった. g/L)あるいは8-bromo-cAMP(1mM)は24時間の添加でhCG,hCG-およびPの分泌を統計的有意差をもって促進した.その分泌促進作用はRAより著明であった.RAとcholera toxinの両者の添加実験により,hCGおよびhCG-分泌は相乗的に促進された.また,P分泌は相加的に促進された.細胞内cAMP含量はRA添加により促進されなかった(表1).RA(10-10-5×10-6M)は細胞増殖あるいは細胞蛋白含量に影響を与えなかった. 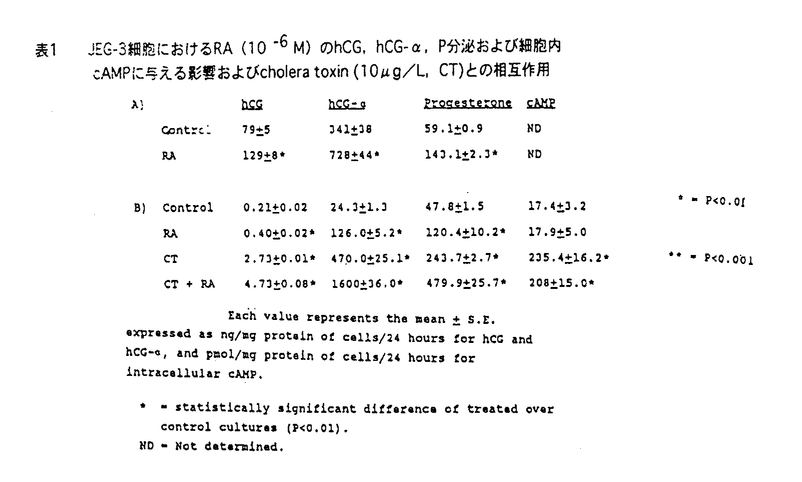 表1 JEG-3細胞におけるRA(10-6M)のhCG,hCG-,P分泌および細胞内cAMPに与える影響およびcholera toxin(10g/L,CT)との相互作用2.Jar細胞におけるRAの影響 表1 JEG-3細胞におけるRA(10-6M)のhCG,hCG-,P分泌および細胞内cAMPに与える影響およびcholera toxin(10g/L,CT)との相互作用2.Jar細胞におけるRAの影響 4日間の添加でRAは濃度依存的にhCGおよびhCG-の分泌を促進した.10-6MのRAは48時間後に統計的に有意なhCGおよびhCG-の分泌促進作用を示した.RAはPの分泌にはなんらの影響を与えなかった.また,細胞内cAMP含量もRAにより影響されなかった.一方,cholera toxin(10g/L)はhCG,hCG-,P分泌を促進し,細胞内cAMP含量を増加させた.RAとcholera toxinの同時添加により,hCGおよびhCG-分泌を相加的あるいはそれ以上に促進したが,P分泌あるいは細胞内cAMP含量はcholera toxin単独添加による以上には影響されなっかた(図1). 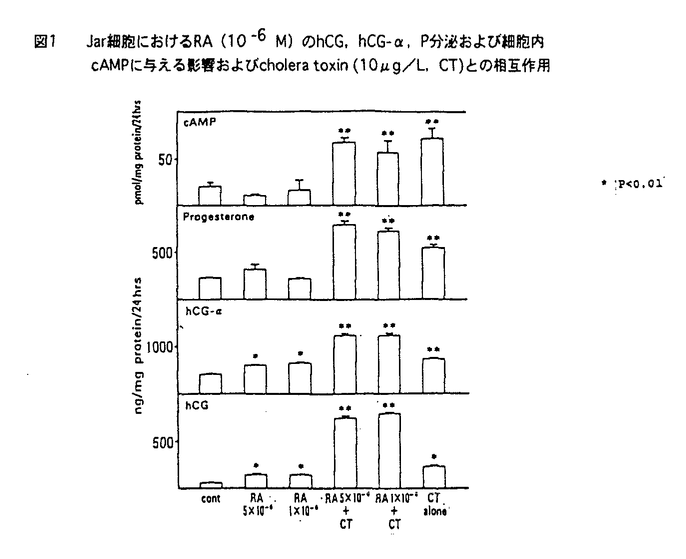 図1 Jar細胞におけるRA(10-6M)のhCG,hCG-,P分泌および細胞内cAMPに与える影響およびcholera toxin(10g/L,CT)との相互作用3.BeWo細胞におけるRAの影響 図1 Jar細胞におけるRA(10-6M)のhCG,hCG-,P分泌および細胞内cAMPに与える影響およびcholera toxin(10g/L,CT)との相互作用3.BeWo細胞におけるRAの影響 RAは濃度依存的にhCGおよびhCG-分泌を促進した.統計的に有意の促進作用を示すRAの最低濃度は10-8Mであった.P分泌もRAにより促進されたが,統計的有意差を示すのに必要なRAの濃度は10-6Mであった(表2).cholera toxin(10g/L)はJarあるいはJEG-3細胞と同様にhCG,hCG-およびP分泌を促進した.cholera toxin(10g/L)とRAの同時添加によりhCG,hCG-およびP分泌は相加的に促進した. 4.正常絨毛細胞におけるRAの影響 妊娠初期および末期胎盤より得られたcytotrophoblastの培養系においてRAはhCG,hCG-およびP分泌になんらの影響を与えなかった. 考察 (1)RAのhCG,hCG-分泌促進作用は今までJEG-3細胞でのみ報告されていた.今回3種類の絨毛癌細胞株で確認されたことより絨毛癌細胞においてはRAの作用は一般的であると考えられた.(2)RAのP分泌促進作用は2種類の絨毛癌細胞でみられたが,他の1種類の絨毛癌細胞ではみられなかったこと,および有意の分泌促進に要するRAの濃度がhCG,hCG-に比べ高いことより,絨毛癌細胞においてはRAのP分泌に対する作用は一定でないことが示された.(3)RAはcAMPを直接には介さない経路でhCG,hCG-およびP分泌を促進することが明らかとなった.(4)RAのhCG,hCG-,P分泌促進作用は正常の絨毛細胞では認められなかったことより,RAの胎盤ホルモンの分泌をmodulateする能力は絨毛細胞の癌化により獲得された生物学的性格であることが示された.(5)以上より,絨毛細胞の癌化とRAに対する反応性の獲得との間に関連性があることが示唆された. |
| 審査要旨 | | 絨毛癌細胞および正常絨毛細胞はホルモンを産生するが、本研究はこれらの細胞の内分泌機能が,分化誘導因子であるレチノール酸により調節されているかを,ヒト絨毛癌細胞株および正常絨毛の培養系を用いて解析したもので以下の結果を得ている. 2.同様に3種類の細胞株において,hCG,hCG-分泌促進作用を有することで知られているcAMP pathwayを刺激する8-bromo-cAMP/cholera toxinとレチノール酸との同時添加実験では,hCG,hCG-分泌は相乗的,あるいは相加的に促進された.また,レチノール酸単独の添加では細胞内cAMP含量は影響されなかったことより,レチノール酸のhCG,hCG-分泌促進作用はcAMPを直接増加させることによるものではないことが明らかとなった. 3.一方,これら絨毛癌細胞株において,レチノール酸のプロゲステロン分泌に与える影響に関しては一定の傾向が見られなかった.すなわち,JEG-3およびBeWo細胞においてはレチノール酸はプロゲステロン分泌を促進したが,Jar細胞においては影響を与えなかった. 4.妊娠末期および妊娠初期の正常絨毛細胞の初代培養による実験系においては,培養の過程で絨毛細胞は未分化な栄養膜細胞(cytotrophoblast)から分化した栄養膜合胞体(syncytiotrophoblast)へと形態的、機能的に分化する.この絨毛細胞のin vitro分化モデルの実験系において,レチノール酸を,絨毛細胞が未分化な栄養膜細胞である時期あるいは栄養膜合胞体に分化した時期に添加してもhCG,hCG-およびプロゲステロン分泌になんらの影響を与えなかった.また,栄養膜細胞から栄養膜合胞体への形態的分化にも影響を与えなかった. 以上,本論文は分化誘導因子であるレチノール酸の絨毛癌細胞および正常絨毛細胞の内分泌機能に与える影響を詳細に検討していて,レチノール酸の内分泌調節作用は癌化した絨毛細胞では見られるが,正常絨毛細胞では見られないことを明らかにした.これらの結果は絨毛細胞の癌化とレチノール酸に対する反応性の獲得との間に関連性があることを示唆した点でユニークであり,従来明らかにされていなかったレチノール酸の絨毛糸細胞における細胞機能の調節作用に関して重要な貢献をなすと考えられ,学位の授与に値するものと考えられる. |