本研究は、マクロファージスカベンジャー受容体(MSR)欠損マウスを用いてヒトMSR蛋白に対する単クローン抗体を樹立し、得られた抗体を用いてヒト脳におけるMSRの発現分布を免疫組織学的に明らかにしたもので、下記の結果を得ている。 1.通常の方法では作成困難であったヒトMSR蛋白に対する特異的な単クローン抗体を、MSR欠損マウスを用いて樹立した。抗原には、大腸菌またはバキュロウイルスを用いて作成したヒトI型MSR組換え蛋白を用いた。ELISAによるスクリーニングのあと、Western blotting及びMSR発現組織での免疫組織学的検討により抗体の特異性を確認した。種をこえて保存される蛋白に対する抗体作成には、その遺伝子が欠損したマウスを用いて免疫寛容類似の状態を擬似的になくすことが有用であることが示唆された。 2.抗ヒトMSR抗体を用いたヒト脳組織の検討で、病変の有無に関わらず脳微小血管周囲にMSR陽性細胞が一貫して観察された。この細胞はCD68陽性であることからマクロファージ系細胞であり、脳微小血管周囲にあって紡錐形の胞体を持ち相互に連絡するという形態・局在両面の特徴から間藤細胞と同定された。本研究により、MSR陽性の間藤細胞がヒト脳内にも存在することが蛋白レベルで初めて証明された。動物での検討から間藤細胞は血液脳関門の一部をなし、老廃物を除去する機能を持つことが知られている。間藤細胞が微小血管周囲をネットワークをなして取り囲むことが証明され、間藤細胞の機能にMSRが重要な役割を果たすことが示唆された。 3.通常の神経病理学的検索で病変をみとめない脳組織においては、ミクログリアでのMSR発現は検出感度以下に抑制されていた。 4.Alzheimer病におけるアミロイド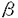 蛋白の沈着部位や、多発性硬化症の脱髄巣など様々な脳病変に伴ってミクログリアが反応性に活性化すると、その一部にMSRが発現誘導された。脳虚血病巣ではMSRを発現している反応性ミクログリアが非常に多く観察された。ミクログリアは障害組織の除去と修復をする働きをもつことが知られ、ミクログリアが変性神経細胞の除去にあたってMSRを介した機構を利用していることが考えられた。 蛋白の沈着部位や、多発性硬化症の脱髄巣など様々な脳病変に伴ってミクログリアが反応性に活性化すると、その一部にMSRが発現誘導された。脳虚血病巣ではMSRを発現している反応性ミクログリアが非常に多く観察された。ミクログリアは障害組織の除去と修復をする働きをもつことが知られ、ミクログリアが変性神経細胞の除去にあたってMSRを介した機構を利用していることが考えられた。 5.ミクログリアによるMSRの発現は疾患特異的ではないことから、様々な病変に共通してMSRが機能することが示唆された。また反応性ミクログリアのすべてがMSR陽性ではなく、MSR特異な発現調節機構の存在が想定された。 6.Alzheimer病症例の中で、血管壁にアミロイド蛋白が沈着するアミロイドアンギオパチーを呈した症例では、アミロイド沈着血管にMSRを強く発現した間藤細胞が観察された。アミロイド蛋白の沈着は産生と除去のバランスに影響されると考えられるが、その過程にミクログリアだけでなく間藤細胞も関わることが示唆された。 以上のように、本論文はMSR欠損マウスを用いてMSRに対する単クローン抗体を作成し、その抗体を用いてヒト脳におけるMSRの免疫組織学的検討を行い、生理的条件下で間藤細胞がMSRを介して老廃物を除去している可能性を示したものである。さらに病的過程においてミクログリアにもMSRの発現がみられることを見出している。これらの結果は、生理的及び病的条件下でのヒト脳内の老廃物処理機構に関して新らたな知見を提供するものであり、学位の授与に値すると認められる。 | 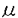 m厚の切片を作成した。脳組織は剖検時に厚さ5mm程度の小さなブロックとして切り出し、4%パラホルムアルデヒドで48時間固定し、一部は70%エタノールで24時間固定した。15%ショ糖溶液中で凍結保護したのち凍結ミクロトームで30
m厚の切片を作成した。脳組織は剖検時に厚さ5mm程度の小さなブロックとして切り出し、4%パラホルムアルデヒドで48時間固定し、一部は70%エタノールで24時間固定した。15%ショ糖溶液中で凍結保護したのち凍結ミクロトームで30