造血前駆細胞から異なる樹状細胞群への分化経路を想定し、マウス骨髄由来Lin-c-kit+造血前駆細胞を培養下にてGM-SCF、SCF、TNF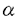 で刺激し、CD11bとCD11cの発現を指標に樹状細胞の発生・分化を細胞生物学的に考察し、下記の如き結果となった。 で刺激し、CD11bとCD11cの発現を指標に樹状細胞の発生・分化を細胞生物学的に考察し、下記の如き結果となった。 1、GM-SCF、SCF、TNFで刺激後6日目にLin-c-kit+造血前駆細胞から2つの異なる樹状細胞前駆体である、CD11b-/dullCD11c+とCD11b+hiCD11c+細胞が誘導された。CD11b-/dullCD11c+前駆樹状細胞はCD11b+hiCD11c+前駆樹状細胞に比べ、Ia、CD86、E-カドヘリン、及びDEC-205の発現が高かった。更にCD11b+hiCD11c+前駆樹状細胞では発現の認められないCD40はCD11b-/dullCD11c+前駆樹状細胞では高発現し、この2つの樹状細胞前駆体は数多くの表面マーカーの発現パターンの相違から表現型においても相互に異なる細胞集団と考えられた。 2、CD11b-/dullCD11c+前駆樹状細胞とCD11b+hiCD11c+前駆樹状細胞の両者は更にGM-SCF、及びTNF刺激後10-14日目に表現型上相異なる成熟樹状細胞へ分化した。c-fmsはCD11b+hiCD11c+前駆及び成熟樹状細胞に分化した細胞群に限局して発現したが、CD11b-/dullCD11c+群では検出できなかった。また、CD11b+hiCD11c+前駆及び成熟樹状細胞はCD11b-/dullCD11c+群に比較し、非特異的エステラーゼ活性が認められた。更にCD11b-/dullCD11c+前駆細胞由来成熟樹状細胞では、ランゲルハンス細胞マーカーである、E-カドヘリンが高発現していた。以上、CD11b-/dullCD11c+及びCD11b+hiCD11c+前駆細胞より由来する成熟樹状細胞は表現型上相異なると示唆された。 3、分化過程においてCD11b-/dullCD11c+及びCD11b+hiCD11c+前駆樹状細胞により同種動物由来Tリンパ球増殖誘導能を獲得する一方で、エンドサイトーシス能は著明に減少した。超微細構造所見では、両群の前駆樹状細胞の形態は小型で、大部分は球状であり、成熟化するに連れ、大型化し、細胞質突起の形成及びよく発達した管胞構造、胞体、及び多胞体が観察し得た。これらの結果から、CD11b-/dullCD11c+及びCD11b+hiCD11c+前駆樹状細胞は形態学的にも機能的にも未成熟であり、成熟樹状細胞とは対照的であった。 4.CD11b+hiCD11c+前駆樹状細胞はCD11b-/dullCD11c+前駆樹状細胞と異なり、M-CSF刺激によりマクロファージへ分化した。このマクロファージは細胞内顆粒が豊富で、非特異的エステラーゼ活性が高かった。また、中等度から高度のCD11b発現があり、Ia、CD11c、CD86、あるいはDEC205の発現は認められず、同種リンパ球への増殖刺激能は消失した。これに対して、CD11b-/dullCD11c+前駆樹状細胞はM-CSF存在下でもマクロファージへ分化できず、刺激後3日以内に死滅した。このことから、CD11b+hiCD11c+前駆樹状細胞は、潜在的には2方向への分化能力があり、添加する成長因子に依存し、成熟樹状細胞或いはマクロファージへ分化する一方で、CD11b-/dullCD11c+前駆樹状細胞は成熟樹状細胞への一方向への分化のみ観察された。 これらの結果を総合すると、CD11b-/dullCD11c+、CD11b+hiCD11c+細胞はLin-c-kit+造血前駆細胞に由来する別個の前駆樹状細胞であり、各々は更に別々の経路を辿り成熟樹状細胞へ分化する。マウス造血前駆細胞から培養下で、2つの異なる分化経路が存在することな本論文により確立された。この知見は、樹状細胞分化の機構、MHCクラスII・抗原ペプチド複合体形成後、同複合体の細胞内移動の分子機構並びに多様な免疫応答調節に複数の樹状細胞群が果たす個々の機能を解明するための布石となると思われる。免疫学上での貢献度を勘案すれば、彼の業績は学位授与に値する。 |