細胞間あるいは細胞とそれを取り巻く細胞外マトリックス(ECM)の間の接着は、すべての多細胞生物の発生、生理現象、および病態において重要な役割を担っている。この接着に関与する細胞表面レセプターにはいくつかのファミリー(同族蛋白質群)が同定されているが、中でも主要なファミリーはインテグリンファミリーである。インテグリンは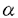 と と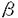 のサブユニットからなるヘテロダイマー構造の細胞表面蛋白質で、ECMを構成する蛋白質や別の細胞表面レセプターと結合する。インテグリン8サブユニットはvサブユニットとのホモロジーにより最近クローニングされた。8サブユニットは1サブユニットとのみヘテロダイマー(81)を形成する。これまでの研究により、81はファイブロネクチン(FN)、ビトロネクチン(VN)、およびテネイシン-C(TN-C)のレセプターとして機能することが示されていた。しかし、生体内における81の機能については全く不明であった。そこで本研究では、このインテグリンの機能について2つの異なる方法を用いて検討した。すなわち、まず、マウスのジーンターゲティング(いわゆるノックアウトマウスの作製)により8サブユニットの遺伝子を欠損させ、発生過程における8蛋白質の欠損の影響を調べた。次に第二の実験として、生体組織上で81リガンドが存在する部位を同定し、未知のリガンドを探索した。その際、高感度で特異的な検出を行うために遺伝子組み換えによって作製した81とアルカリフォスファターゼのキメラ分子をレセプタープローブ(その酵素活性によりリガンド結合が検出できるもの)とする新しい方法を開発して用いた。 のサブユニットからなるヘテロダイマー構造の細胞表面蛋白質で、ECMを構成する蛋白質や別の細胞表面レセプターと結合する。インテグリン8サブユニットはvサブユニットとのホモロジーにより最近クローニングされた。8サブユニットは1サブユニットとのみヘテロダイマー(81)を形成する。これまでの研究により、81はファイブロネクチン(FN)、ビトロネクチン(VN)、およびテネイシン-C(TN-C)のレセプターとして機能することが示されていた。しかし、生体内における81の機能については全く不明であった。そこで本研究では、このインテグリンの機能について2つの異なる方法を用いて検討した。すなわち、まず、マウスのジーンターゲティング(いわゆるノックアウトマウスの作製)により8サブユニットの遺伝子を欠損させ、発生過程における8蛋白質の欠損の影響を調べた。次に第二の実験として、生体組織上で81リガンドが存在する部位を同定し、未知のリガンドを探索した。その際、高感度で特異的な検出を行うために遺伝子組み換えによって作製した81とアルカリフォスファターゼのキメラ分子をレセプタープローブ(その酵素活性によりリガンド結合が検出できるもの)とする新しい方法を開発して用いた。 最初にマウスの発生における81の役割を調べるため、マウスの8サブユニットのcDNAを単離し、これをもとに8欠損マウスを作製した。8遺伝子を変異させたマウスはインテグリン81を発現せず、腎臓の退縮や形成不全がみられ、ほとんどのマウスは生後2日以内に死亡した。他の臓器では出生時までに異常はみられなかった。野生型(正常)マウスの腎臓の形態形成においては、後腎の間葉と尿管上皮との間で相互に交わされる誘導作用が必須である。8欠損マウスでは、尿管芽の成長と分岐、および間葉系細胞の上皮系組織への転換融合が不完全であった。マウス8に対する抗体を用いた免疫組織染色法により正常マウス胚における8の発現分布を調べたところ、8は様々な器官の間葉系細胞で発現していたが、中でも後腎の間葉では、尿管芽との接触によって8の発現が誘導されていることを示唆する発現パターンが観察された。8を発現した間葉系細胞では、続いてその間葉系細胞の上皮系への転換に伴い8の発現は消失するが、成長した尿管上皮と隣り合う位置にある間葉系細胞では、上皮に接する側でのみ8の発現は維持されており、上皮に対する間葉の細胞極性の形成に8の発現が関与することが示唆された。よって、81を介した上皮-間葉間の相互作用が腎臓の形態形成に決定的な役割を果たしていると予想される。8欠損マウスでは、尿管芽と間葉の間の81を介した誘導作用がおきず、上皮-間葉間の細胞極性の形成が異常になるため、腎臓の形態形成が停止するか、あるいは不完全に進行し、腎臓に重篤な欠陥が起こったと考えられる。81を介した上皮-間葉間の相互作用には81のリガンドの存在が必須であるが、既知のリガンド(FN、VN、TN-C)の胎仔の腎臓における発現の時期および局在性は8のそれとは異なっていた。このことは、81の機能の実現にはさらに別のリガンドが関与し、そのリガンドが発生における重要な役割を担うことを示唆している。 次に、生体組織上における未知のリガンドの存在部位を同定するため、インテグリンとアルカリフォスファターゼの性質を合わせ持つキメラ分子(融合蛋白質)を作製した。マウスの8の細胞外ドメインのみからなる蛋白質と、1の細胞外ドメインにアルカリフォスファターゼを融合させた蛋白質を、同時に動物細胞に発現させた。この欠失型インテグリンは可溶性で、キメラのヘテロダイマー(8t1-APと呼ぶ)として分泌され、高純度に精製することができた。精製された8t1-APはリガンド結合能を有し、その活性化およびリガンド特異性は、細胞表面に発現させた81のそれとよく一致していた。リガンド蛋白質をコートしたプラスチックプレートを用いて8t1-APの結合を調べる実験では、8t1-AP分子はFNやVNに結合したが、TN-Cには結合しなかった。前述のようにこれまでの細胞を用いた実験ではTN-Cは81の基質であるとされていたので、この結果は意外であった。その矛盾の原因を探るために、遺伝子工学的に作製した様々なTN-Cの断片を用いて8t1-APの結合を調べた結果、RGD(アルギニンーグリシンーアスパラギン酸)細胞接着ドメインという多くのインテグリンによって認識される共通の配列を含む短い断片にのみ、顕著に結合することが判明した。この結合はRGD配列を含むペプチドにより完全に阻害された。またTN-CのRGDを含む断片でも長さの長いものは8t1-APに結合しなかった。このことは、TN-CのRGD部位は81にとっては隠れた(cryptic)結合部位で、断片化などにより露出されてはじめて結合できるようになるものであることを示唆している。これまで81とリガンドの結合を検出する研究において、細胞を用いた方法では、他の接着レセプターや細胞内情報伝達系の影響を受け、細胞膜上のインテグリンを精製して用いる方法では界面活性剤の影響を免れなかった。8t1-AP分子を用いた方法は、従来の方法に比べて高感度に生理的条件下でリガンド結合を検出できる優れた方法であると考えられる。 さらにこの8t1-APをプローブとして、マウス胚の組織切片やホールマウント(全形のまま固定した)器官を染色し、生体組織上の81リガンドの分布を調べた。胎仔の骨および腎臓における81リガンドの分布は、免疫組織染色やin situハイブリダイゼーションによるオステオポンチン(OPN)の発現分布と重なっていた。OPNはRGD配列を含むECM蛋白質の一つで、胚発生、創傷治癒、癌化などで発現されることが報告されている。この8t1-APによる染色はRGD配列を含むペプチドにより完全に阻害された。興味深いことに、腎臓では、81リガンドの分布は8サブユニットの発現の分布と相補的な位置関係にあり、8が間葉系細胞で発現されるのに対し、81リガンドは尿管上皮細胞の外側表面に局在していた。この分布様式から、この81リガンドは腎臓の形態形成に関与する調節因子として有力な候補であると考えられる。胎仔腎臓の抽出物をゲル電気泳動法により複数の蛋白質バンドに分離し、それに対する8t1-APの結合を調べたところ、既知のリガンドであるFNやVNと同じ分子量のバンドのほかに、複数の未知の蛋白質のバンドへの結合が検出されたが、その中にはOPNと同じ分子量のものが含まれていた。精製したOPNに対しても8t1-APは結合し、その結合はRGDペプチドにより完全に阻害された。また、遺伝子導入により81を発現させた浮遊細胞は、本来接着しなかったOPNに接着し、細胞表面の81もOPNに結合することが明らかになった。最近、OPNに対する抗体およびRGDペプチドが器官培養した腎臓の形態形成を阻害することが他の研究者らによって報告された。その結果と合わせて考えると、本研究の結果は、OPNとインテグリン81の相互作用は腎臓や他の組織の形態形成を調節していることを示唆している。また、本研究における8t1-APの結合実験において検出されたこれまで未同定の蛋白質が腎臓における主要な81リガンドとして機能していることも予想される。 以上、本研究は腎臓の形態形成においてインテグリン81が上皮-間葉相互作用を調節しているという遺伝学的証拠を示した。また、OPNがインテグリン81のリガンドの一つであることを明らかにし、このインテグリンとOPNの相互作用がマウス発生において何らかの役割を担っていることを示唆した。さらに、本研究で開発したインテグリンとアルカリフォスファターゼとのキメラ分子が、様々な生理現象や発生において重要なインテグリン(または他の細胞接着レセプター)とそのリガンドの相互作用を探索する道具として、有効であることを示した。 | 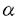 8
8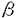 1の役割
1の役割