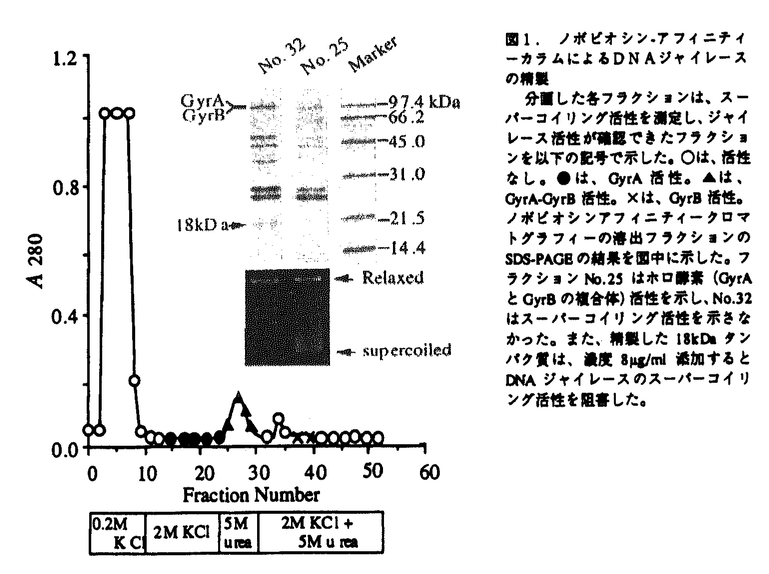
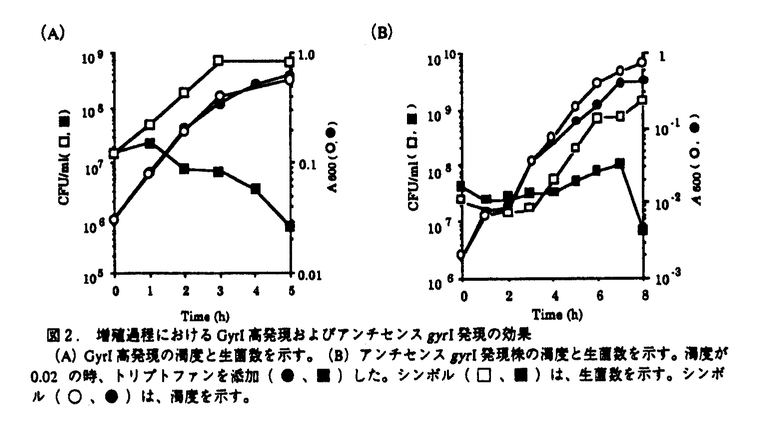
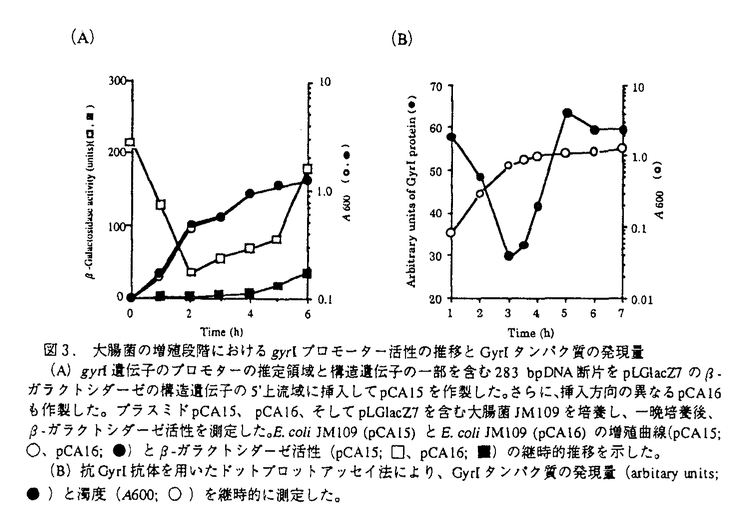
本研究は、大腸菌のDNAスーパーコイリングの制御機構を解明することを目的として、DNAジャイレースのスーパーコイリング活性を阻害するタンパク質(GyrI)を見出し、本タンパク質をコードする遺伝子(gyrI)のクローニングおよび塩基配列の解析、他菌種でのgyrI相同遺伝子の存在有無、さらにGyrIタンパク質の生理的作用の解析を試みたものであり、下記の結果を得ている。 1.ノボビオシン-アフィニティークロマトグラフィーを用いた大腸菌DNAジャイレースの精製過程で、各フラクションのスーパーコイリング活性の測定およびSDS-PAGE電気泳動による解析を行ったところ、DNAジャイレースのホロ酵素を含むにもかかわらず、スーパーコイリング活性を示さないフラクションが存在し、このフラクションには18kDaのタンパク質が含まれていることを見出した。精製した本タンパク質(GyrI)は、スーパーコイリング活性を阻害し、DNAポリメラーゼやテロメラーゼの他酵素の活性に対して阻害作用は認められなかったことから、DNAジャイレースを特異的に阻害する可能性が示唆された。 2.GyrIタンパク質は、N末端アミノ酸配列をもとにホモロジー検索を行ったところ、大腸菌K-12株の染色体DNAの仮想産物であるYeeBと一致した。yeeB遺伝子の翻訳領域前後の塩基配列をもとに、大腸菌gyrI遺伝子をクローニングし、塩基配列を決定した結果、gyrI遺伝子のプロモーターと考えられる-10、-35領域の典型的な配列が示された。また、モチーフ検索の結果、カゼインキナーゼIIのリン酸化部位、プロテインキナーゼCのリン酸化部位、N-ミリストイル化部位のモチーフを有していることが示された。 3.gyrI遺伝子をプローブに用いて、さまざまな細菌の染色体DNAとサザンハイブリダイゼーションを行った結果、Escherichia coli KL16、E.coli ML-1410、E.coli ATCC2592、E.coli KC14、E.coli K-12、Shigella boydii IID627、Shigella dysenteriae IID633、Shigella sonnei TRRL10805にgyrIと高い相同性を示すDNA断片が見出された。また、Citrobacter freundii IID976には中程度の相同性を示すDNA断片が見出された。さらに、Pseudomonas aeruginosa ATCC27853、Baciiius subtilis ATCC6633、Enterococcus faecalis ATCC29212、Staphylococcus aureus RN450に低い相同性を示すDNA断片が見出された。 4.赤痢菌から大腸菌gyrI遺伝子と強くハイブリダイズするDNA断片をクローニングし、塩基配列を解析した結果、大腸菌と赤痢菌のgyrI遺伝子の塩基配列は98%、アミノ酸配列は97%の高い相同性が示された。また、GyrI相同タンパク質が他菌種に存在するかどうかホモロジー検索を行った結果、Bacillus subtilisのyosT遺伝子の仮想産物に対して、アミノ酸配列で22.01%の相同性を示した。さらに、S.typhimuriumにも73.3%の高い相同性を示すタンパク質が存在していることが示された。GyrIは、大腸菌以外の他菌種にもその相同タンパク質が存在する可能性が示された。 5.GyrIをコードするDNAを大腸菌からクローン化し、このDNAを適当なプラスミドに連結し、大腸菌に形質転換してGyrI高発現株を取得した。本発現株から精製したGyrIタンパク質は、DNAジャイレースのスーパーコイリング活性を6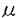 g/mlで阻害した。この濃度は、大腸菌KL16株から精製したGyrIによる阻害濃度とほぼ同じ値を示し、組換えタンパク質は、スーパーコイリング活性阻害能を保持していることが示された。 g/mlで阻害した。この濃度は、大腸菌KL16株から精製したGyrIによる阻害濃度とほぼ同じ値を示し、組換えタンパク質は、スーパーコイリング活性阻害能を保持していることが示された。 6.GyrI高発現株およびアンチセンスRNA発現株を用いて、GyrIタンパク質の機能を調べたところ、両発現株は、増殖を抑制し生菌数を減少させた。この時、大腸菌の形態は、繊維状になり無核細胞や伸長した細胞内に核様体が1つといった核の複製異常が観察された。GyrIはin vivoでもDNAジャイレースの活性を阻害していることが示された。 7. -ガラクトシダーゼ活性を指標としてgyrI遺伝子のプロモーター活性を測定した結果、-ガラクトシダーゼ活性は、対数増殖後期から増加することが示された。また、GyrIのポリクロナル抗体を用いたドットブロットアッセイを行った結果、GyrIタンパク質の発現量はプロモーター活性の推移に連動し、対数増殖後期から増加することが示された。 -ガラクトシダーゼ活性を指標としてgyrI遺伝子のプロモーター活性を測定した結果、-ガラクトシダーゼ活性は、対数増殖後期から増加することが示された。また、GyrIのポリクロナル抗体を用いたドットブロットアッセイを行った結果、GyrIタンパク質の発現量はプロモーター活性の推移に連動し、対数増殖後期から増加することが示された。 以上、本論文では、GyrIタンパク質がDNAジャイレースのスーパーコイリング活性を阻害し、GyrI高発現株やアンチセンスRNA発現株、さらにGyrIタンパク質の発現量の解析から、DNAジャイレースの活性制御に関与していることを明らかにした。本研究は、これまで十分明らかにされていなかったDNAジャイレースの活性制御機構の解明につながり、従来のニューキノロン系抗生物質の耐性菌に対して新しい作用機作の抗生物質を見出すうえでも重要な意義があると考えられ、学位の授与に値するものと考えられる。 |