ストリガ類、アレクトラ類、オロバンケ類に属する寄生雑草は、イネやトウモロコシをはじめとする重要な農作物の根に寄生して水分、栄養分を吸い取り、それらの農作物の収穫量に多大な損害をもたらすために、アフリカ、アジア、そしてアメリカにおいて農民を苦しめ、問題となっている。これらの寄生雑草の種子は、宿主から分泌される物質によってはじめて発芽が誘導されることが知られており、それらの物質の種類・構造に関しては、寄生雑草と宿主の相互認識の観点から長い間関心の高い問題として多くの研究が行われてきた。 1966年に非寄主植物の棉(Gossypium hirsutum)の根の抽出物からストリゴール(1)が単離された。ストリゴールはストリガ類に対して非常に高い活性を示し、10-15Mという極めて低い濃度においても種子の発芽を誘導することが報告されており、その構造は誘導体のX線結晶構造解析により絶対配置まで決定されている。現在までにストリゴール及びその類縁体の合成が数多く検討され、各部の構造が活性に及ぼす影響について精力的に調べられている。 ストリゴールの単離以降も宿主からの発芽誘因物質の検索は続けられ、1992年にソルゴー(Sorghum bicolor)からソルゴラクトンが、同じく1992年にササゲ(Vigna unguiculata)からアレクトロールが、1998年にはアカクローバー(Trifolium pratense)からオロバンコールが単離され、限られたスペクトルの分析からそれぞれ図に示した推定構造が提出された。これらの化合物は類似の骨格を有していることから、総称としてストリゴラクトンと名付けられている。 ストリゴラクトン類は天然から微量しか得られず、しかも単離過程で不安定であるために精製が難しく、したがって構造解析についてはストリゴール以外は完全ではない。そこで本論文では、提唱されている構造を有する化合物を合成し、天然物との物性・生物活性の比較を行うことにより、天然物の構造を確認することができると考え、また発芽誘因物質の構造活性相関に関しても有用な知見が得られることを期待し、ストリゴラクトン類の合成を検討した。  第一章:(±)-ソルゴラクトンとその立体異性体の合成 原料の5から6段階で誘導したジケトカルボン酸6のナトリウム塩のLuche還元で得られるヒドロキシラクトン7の水酸基の臭素化、還元的脱臭素化により、無置換三環性ラクトンの約1:1のジアステレオマー混合物 [(±)-9,(±)-10]へと導いた。この混合物は中圧液体クロマトグラフィー(Lobar LiChroprep Si60)により分離することができ、結晶した方のジアステレオマー(±)-10のX線結晶構造解析により相対配置を決定することができた。続いて、油状の(±)-9をホルミル化した後、別途合成したブロモブテノリド(±)-13と縮合させ、シリカゲルクロマトグラフィーによる分離精製を経て、2種のジアステレオマー(±)-2a,(±)-2bをそれぞれ無色結晶として得ることに成功した。両者のスペクトルはほぼ同一で区別がつかなかったが、極性が低く、融点が127-129℃の結晶についてX線構造解析を行い、目的の構造の(±)-2aであることを明らかにした。同様にして、(±)-10からも(±)-2c、(±)-2dを合成した。 Si60)により分離することができ、結晶した方のジアステレオマー(±)-10のX線結晶構造解析により相対配置を決定することができた。続いて、油状の(±)-9をホルミル化した後、別途合成したブロモブテノリド(±)-13と縮合させ、シリカゲルクロマトグラフィーによる分離精製を経て、2種のジアステレオマー(±)-2a,(±)-2bをそれぞれ無色結晶として得ることに成功した。両者のスペクトルはほぼ同一で区別がつかなかったが、極性が低く、融点が127-129℃の結晶についてX線構造解析を行い、目的の構造の(±)-2aであることを明らかにした。同様にして、(±)-10からも(±)-2c、(±)-2dを合成した。  第二章:(+)-ソルゴラクトンの合成 (S)-シトロネラールを出発原料とし、既知の5段階によりヒドロキシエステル14へと誘導した。14をヨウ化物へと変換した後、三重結合の導入を行い、カルボニル基の 位にフェニルセレニル基を導入してから分子内でラジカル環化反応を行わせることにより、若干の不純物を含む環外アルケン17を得ることに成功した。これと過臭化臭化ピリジニウムを反応させて得られるジブロミド18から3段階で三環性ラクトンのジアステレオマー混合物へと変換し、中圧液体クロマトグラフィーにより9を分離し、(+)-2aへと導くことができた。 位にフェニルセレニル基を導入してから分子内でラジカル環化反応を行わせることにより、若干の不純物を含む環外アルケン17を得ることに成功した。これと過臭化臭化ピリジニウムを反応させて得られるジブロミド18から3段階で三環性ラクトンのジアステレオマー混合物へと変換し、中圧液体クロマトグラフィーにより9を分離し、(+)-2aへと導くことができた。 ソルゴラクトン天然物の構造は、合成したラセミ体及び光学活性体の各種スペクトル分析(NMR,IR,CD)から推定構造にほぼ間違いのないことが示されたが、天然物と合成品で1H-NMRスペクトルに微妙な違いもみられるために最終的な結論はソルゴラクトンの再単離に委ねることとした。また、ラセミ体と光学活性体をあわせた8サンプルについて、ヤセウツボ(Orobanche minor)種子を用いた生物活性試験を実施し、すべての異性体に高い発芽誘導活性があることを見いだした。 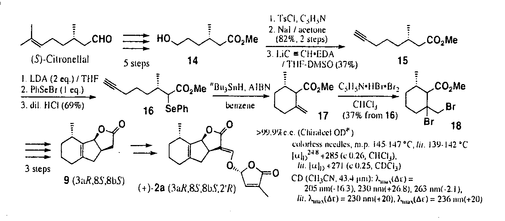 第三章:アレクトロール推定構造を有する化合物とその関連化合物のラセミ体合成 シトラールから文献既知の10段階で(±)-20を大量に合成し、m-クロロ過安息香酸酸化後、エポキシド(±)-21、(±)-22を得た。(±)-21をアルミニウムトリイソプロポキシドで処理することにより、アルコール(±)-23が得られたが、大部分はA環の方に二重結合の入った異性体(±)-24に変換された。(±)-22については、同様の処理をしたところ、ラクトン環の再構築を必要としたが、収率良く(±)-25へと変換させることができた。(±)-25からは、これまでと同様にして(±)-3aと(±)-3bへと変換し、これらと関連中間体について、天然物との詳細な物性比較を行うことより、提唱されていた構造が誤りであることを明らかにすることができた。ストリゴラクトン類は互いに良く似た構造で構造の同定が困難であるため、このような結果は推定構造を有する化合物の合成によってはじめて判明したものである。  また、同様の骨格を有する26をアレクトロールの候補化合物として新たに考え、(±)-20からの(±)-26a、(±)-26bの合成を行ったが、天然物の1H-NMRスペクトルとは一致しなかった。  第四章:オロバンコール推定構造を有する化合物のラセミ体合成と構造検討 , -不飽和ケトン(±)-28のトシルヒドラゾンをシアノ水素化ホウ素ナトリウムを用いて還元し、(±)-31と(±)-32を得た。(±)-32からは第三章と同様の手法により、(±)-35を経て(±)-4a、(±)-4bへと変換し、X線構造解析により両者の構造を決定した。また光延反応を利用して(±)-35の水酸基を反転させた(±)-36からも(±)-4c、(±)-4dを合成した。同様にして(±)-32からも4異性体(±)-4e〜(±)-4hを合成し、8種類全ての立体異性体をラセミ体として得ることができた。 -不飽和ケトン(±)-28のトシルヒドラゾンをシアノ水素化ホウ素ナトリウムを用いて還元し、(±)-31と(±)-32を得た。(±)-32からは第三章と同様の手法により、(±)-35を経て(±)-4a、(±)-4bへと変換し、X線構造解析により両者の構造を決定した。また光延反応を利用して(±)-35の水酸基を反転させた(±)-36からも(±)-4c、(±)-4dを合成した。同様にして(±)-32からも4異性体(±)-4e〜(±)-4hを合成し、8種類全ての立体異性体をラセミ体として得ることができた。 次いで、これらのTMS誘導体のGC-MS分析により、提唱されていた構造が誤りであることを明らかにし、さらに関連化合物の分析を進めた結果、GC-MS分析における保持時間、開裂様式の比較及びこれまでの一連の検討結果から、オロバンコール天然物の構造が(±)-26aである可能性が非常に高いことを示すことができた。また、(±)-26a、(±)-26bが高いヤセウツボ種子発芽誘導活性を有していることも確認することができた。  以上、本論文では、寄生雑草種子発芽誘因物質ストリゴラクトン類3種の合成を行い、それらの構造と生物活性について新しい有用な知見を得るとともに、天然から微量しか得られない生体制御物質の構造解析における有機合成の有用性を示すことができた。 |