学位論文要旨
| No | 214120 | |
| 著者(漢字) | 関根,信夫 | |
| 著者(英字) | ||
| 著者(カナ) | セキネ,ノブオ | |
| 標題(和) | インスリン分泌細胞の増殖と機能の制御に関与する因子とその情報伝達 | |
| 標題(洋) | Factors and Their Signaling Mechanisms Involved in the Regulation of Growth and the Function of Insulin-Secreting Cells | |
| 報告番号 | 214120 | |
| 報告番号 | 乙14120 | |
| 学位授与日 | 1999.01.27 | |
| 学位種別 | 論文博士 | |
| 学位種類 | 博士(医学) | |
| 学位記番号 | 第14120号 | |
| 研究科 | ||
| 専攻 | ||
| 論文審査委員 | ||
| 内容要旨 | 糖尿病における糖代謝異常の成因としてインスリン分泌障害は最も重要なものであり,全ての糖尿病患者に量的または質的な膵 細胞増殖・機能の検討:細胞増殖・生存率は細胞数算定,MTTアッセイ,[3H]チミジン摂取率(DNA合成)測定により評価した.インスリン分泌能についてはブドウ糖をはじめとする各種刺激因子に対する反応性を静置実験で検討した.細胞内代謝は,インスリン分泌と平行してMTTアッセイで評価するか,14C標識ブドウ糖・ピルビン酸を用いて測定した.細胞内Ca2+([Ca2+]i)の測定:Ca2+感受性蛍光指示薬fura-2を細胞に取り込ませ,単一細胞レベルまたは顕微鏡画像処理により複数の細胞における[Ca2+]iの変化を観察した.細胞膜電位の測定:蛍光指示薬ビスオキソノールまたは電気生理学的手法(パッチクランプ法)により検討した.免疫沈降・ウェスタンブロット:蛋白チロシンリン酸化を検討するため,細胞質蛋白を当該の特異的抗体で免疫沈降した後電気泳動し,抗リン酸化チロシン抗体でイミュノブロットを行った.ノーザンブロット:細胞よりRNAを抽出し,当該の32P標識プローブを用いて行った.MAPK/MEKアッセイ:細胞質蛋白を44-kDa MAPKまたはMEK-1特異的抗体で免疫沈降した後,MAPK活性はミエリン塩基性蛋白のリン酸化能を,MEK-活性はリコンビナントMAPK活性化能を指標として評価した.ゲルシフトアッセイ:各転写因子の特異的DNA結合部位に相当する合成オリゴヌクレオチドを32P標識プローブとして用い,核蛋白との結合を電気泳動度により評価した. I-1.SFMにおけるINS-1細胞の増殖に影響する因子:SFM中に培養されたINS-1細胞は血清添加とほぼ同様の生存率・増殖曲線を示した.SFMに添加されているPRLを除くとINS-1細胞によるチミジン摂取率が半減したことから,PRLがSFMの中で最大の増殖刺激活性を有することが確認された.15mMブドウ糖では3mMに比してDNA合成は約6倍促進された.またcAMPを上昇させる因子はブドウ糖による増殖刺激作用を阻害した.一方,受容体チロシンキナーゼを介して作用する増殖因子(EGF,PDGF,bFGF,NGF)に関しては顕著な効果が見られなかった. I-2.GH・PRLの生理活性:SFM中でGHおよびPRLはINS-1細胞の増殖,生存,代謝活性を刺激した.細胞内インスリン含量は非添加群で著明に低下したが,GH・PRL添加によりその低下が阻止された.またGH,PRLともに1pM-1nMの生理的濃度で用量依存性にDNA合成を刺激し,両者の活性はほぼ同等であった. I-3.IFN- I-4.INS-1細胞のインスリン分泌能を保持する因子:SFM内に3日間培養されたINS-1細胞は栄養素刺激によるインスリン分泌の著明な低下を示したが,高濃度K+によるそれは障害されず,SFM中にはINS-1細胞の代謝活性ひいてはインスリン分泌能を保持する何らかの因子が欠如しているものと考えられ,検討を加えた.まず血清再添加により分泌能はほぼ完全に回復した.次にSFM内に各種因子を補充して分泌能低下が阻止されうるか検討したが,明らかな効果を示したものはなかった.この分泌障害の機序を細胞内代謝,膜脱分極,[Ca2+]iの変化について検討したところ,栄養素の酸化的代謝が著しく低下し,それにより後2者の刺激も障害されていた.最後にラット膵島または II-1.GH・PRLの情報伝達:まずGH・PRLによる[Ca2+]i上昇作用について検討した.この作用は細胞外Ca2+をEGTAでキレートしたり,L型Ca2+チャネル阻害薬やジアゾキシドを添加することにより抑制されたことから,電位依存性Ca2+チャネルを通してのCa2+流入によるものと考えられた.またAキナーゼ阻害薬Rp-cAMPSによっても[Ca2+]i上昇は阻害されたためcAMPの関与も示唆されたが,実際に細胞内cAMPの上昇は検出できなかった.さらにGH・PRLによる蛋白チロシンリン酸化を検討した.両ホルモンはJAK2キナーゼおよびSTAT5のリン酸化を促進し,STAT5の活性化はゲルシフトアッセイでも確認された.最後にGHの細胞増殖刺激作用に関与する情報伝達を明らかにするため,各種阻害薬のGH作用に及ぼす影響を検討した.その結果,GHの[Ca2+]i上昇作用を阻害するベラパミル・ジアゾキシドはDNA合成に影響せず,チロシンキナーゼ阻害薬ラベンダスチンAおよびRp-cAMPSによって抑制が認められた. II-2.IFN- II-3.INS-1細胞の増殖・機能の制御におけるMAPKの役割:まず各種インスリン分泌刺激因子によるMAPKの活性化について検討した.ブドウ糖刺激はMAPK活性の持続的上昇をもたらした.cAMPを上昇させる因子はいずれも単独での作用は弱いが,ブドウ糖によるMAPK活性化を著明に増強した.これらMAPK活性化のパターンや用量反応性は,同時に測定されたインスリン分泌のそれとよく一致した.またCキナーゼ活性化によりインスリン分泌を刺激するPMAや高濃度K+によるCa2+流入もMAPKを活性化した.他方,INS-1細胞の増殖を刺激するGHはMAPKの活性に影響しなかったのに対し,NGFはINS-1細胞において強力なMAPKの活性化因子であり,インスリン分泌刺激作用を有することが確認された.これらINS-1細胞におけるMAPK活性化因子は[Ca2+]i上昇作用をも有するため,両作用の関連について検討した.まずブドウ糖によるMAPK活性化およびcAMPによるその増強はEGTAやベラパミルによって70-90%阻害され,これらの機序としてCa2+流入の必要性が示された.一方,PMAやNGFによるMAPK活性化には[Ca2+]i上昇は必須でないと考えられた.またブドウ糖,CPT-cAMP,高濃度K+はいずれもMAPKの上流に存在するMEK1を活性化し,これもEGTA処理により阻害されたことから,これらの因子は[Ca2+]i上昇を介してMEK1,さらにはMAPKを活性化するものと考えられた.最後にMAPK活性化因子による早期応答遺伝子の誘導について検討したところ,[Ca2+]i上昇を介するMAPK活性化にはjunBの誘導がよい相関を示し,NGFは特にzif268を顕著に誘導した. (1) | |
| 審査要旨 | 本研究はインスリン分泌細胞の増殖と機能の制御に関与すると考えられる種々の因子とその細胞内情報伝達機序を明らかにするため,分化した形質を有する培養ラットインスリノーマ細胞INS-1を用いた系を確立し,広く検討を行ったものである.その中で,とくにインスリン分泌細胞の増殖・機能に対し促進的に作用する成長ホルモン(GH)・プロラクチン(PRL),および抑制的に作用するサイトカインであるインターフェロン- 1.INS-1細胞は種々の生理的膵 2.無血清培地中に培養されたINS-1細胞は血清添加時同様の増殖能を示したが,ミトコンドリア代謝の低下を主要因とするインスリン分泌障害を呈した.この障害は 3.GH・PRLはINS-1細胞の増殖およびインスリン合成を刺激した.両ホルモンとも,細胞膜脱分極に引き続き電位依存性Ca2+チャネルを通して細胞外Ca2+の流入を促進することにより細胞内Ca2+を上昇させるが,この作用は増殖刺激には必須ではなかった.これに対し,チロシンキナーゼJAK2とそれに続く転写因子STAT5の活性化が,GH・PRLによる増殖刺激作用の情報伝達機序として主要なものであることが示唆された. 4.IFN- 5.[Ca2+]i上昇,cAMP上昇,およびCキナーゼ活性化を介してインスリン分泌を促進する様々な因子が,INS-1細胞においてMAPキナーゼを活性化することが示された.MAPキナーゼの活性化それ自体はインスリン分泌刺激に必ずしも十分ではないが,分泌を促進すべく細胞内の情報伝達系を修飾している可能性が示唆された.一方,INS-1細胞増殖刺激とMAPキナーゼ活性化との間には明確な相関関係は認められなかった. 以上,本論文は優れた膵 | |
| UTokyo Repositoryリンク | http://hdl.handle.net/2261/54097 |
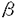 細胞障害が存在するとも言える.従って
細胞障害が存在するとも言える.従って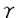 (IFN-
(IFN-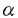 と相乗的に作用して著明な細胞障害性を示した.またIFN-
と相乗的に作用して著明な細胞障害性を示した.またIFN-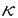 Bを活性化した.興味深いことに,IFN-
Bを活性化した.興味深いことに,IFN-