| 内容要旨 | | 一般に細胞は増殖時には分化した機能は低下すると考えられている。肝においても,70%切除後ラット再生肝でアルブミンmRNAの減少が認められ,肝細胞のDNA合成が最大になる術後24時間目に最低になると報告されている。また,ラットの四塩化炭素肝障害後の肝細胞増殖に際して肝でのアルブミン遺伝子の転写が低下し,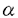 -フェトプロテインの転写と相反して変動することが示されている。さらに急性ウイルス性肝炎患者の回復過程では,アルブミンと-フェトプロテインの血中値の相反する変動がみられ,肝再生を促進するグルカゴン-インスリン療法を施行した際にはより顕著になることが観察されている。これらの報告からは,肝細胞固有の機能であるアルブミン合成は肝細胞増殖の際に低下していると考えられる。しかし一方で,部分切除後再生肝において肝重量当たりのアルブミン合成はほとんど変化しないとの報告も認められる。分化した肝にみられる転写因子C/EBPが,再生肝のほぼすべての肝細胞核に発現し,しかもこの発現はDNA合成を行っている肝細胞にもみられるとのデータから,肝細胞は増殖の間も分化した状態を保っているとする報告もある。また,肝細胞のDNA合成を促進する因子として知られているインスリン,グルカゴン,T3,成長ホルモンは,肝細胞の分泌蛋白合成に対しては,インスリン,甲状腺ホルモン,成長ホルモンが蛋白合成を促進し,グルカゴンがこれを低下させることが認められている。Interleukin6は,従来肝に対しては急性相蛋白(フィブリノーゲン,ハプトグロビン等)の産生を増加させ,非急性相蛋白(アルブミン,プレアルブミン等)の産生を低下させる因子として知られていたが,最近interleukin6欠損マウスで肝再生不全が起こり,interleukin6投与により回復することから,肝細胞増殖の必須因子と報告された。肝細胞増殖と肝細胞機能,特に蛋白合成との関係はいまだ明確でない。 -フェトプロテインの転写と相反して変動することが示されている。さらに急性ウイルス性肝炎患者の回復過程では,アルブミンと-フェトプロテインの血中値の相反する変動がみられ,肝再生を促進するグルカゴン-インスリン療法を施行した際にはより顕著になることが観察されている。これらの報告からは,肝細胞固有の機能であるアルブミン合成は肝細胞増殖の際に低下していると考えられる。しかし一方で,部分切除後再生肝において肝重量当たりのアルブミン合成はほとんど変化しないとの報告も認められる。分化した肝にみられる転写因子C/EBPが,再生肝のほぼすべての肝細胞核に発現し,しかもこの発現はDNA合成を行っている肝細胞にもみられるとのデータから,肝細胞は増殖の間も分化した状態を保っているとする報告もある。また,肝細胞のDNA合成を促進する因子として知られているインスリン,グルカゴン,T3,成長ホルモンは,肝細胞の分泌蛋白合成に対しては,インスリン,甲状腺ホルモン,成長ホルモンが蛋白合成を促進し,グルカゴンがこれを低下させることが認められている。Interleukin6は,従来肝に対しては急性相蛋白(フィブリノーゲン,ハプトグロビン等)の産生を増加させ,非急性相蛋白(アルブミン,プレアルブミン等)の産生を低下させる因子として知られていたが,最近interleukin6欠損マウスで肝再生不全が起こり,interleukin6投与により回復することから,肝細胞増殖の必須因子と報告された。肝細胞増殖と肝細胞機能,特に蛋白合成との関係はいまだ明確でない。 近年,肝細胞の最も強力な増殖促進因子のひとつとしてhepatocyte growth factor(HGF)が同定された。慢性肝疾患患者の血中HGF値は高値であり,慢性活動性肝炎患者の肝でHGFが作用していることを示唆する報告もみられる。HGFの生体内での作用とその意義が注目される。 本論文では,HGFの肝細胞分泌蛋白合成に対する作用を肝細胞増殖との関連を含め,in vivo及びin vitroにて検討した。 In vivo実験:5〜6週齢のSprague-Dawley系雄性ラットに,phosphate buffered saline(PBS)に溶解したリコンビナントヒトHGF(rhHGF)100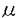 g/kg体重,あるいはPBSを大腿静脈より2時間ごとに6回静注し,初回注射から16,26,48時間後に血液を採取,血清アルブミン及び血漿フィブリノーゲン濃度を測定した。血清アルブミン値はrhHGF投与後26,48時間目にそれぞれ対照の105,117%に上昇した。血漿フィブリノーゲン値も26,48時間目にそれぞれ対照の141,120%に上昇した。 g/kg体重,あるいはPBSを大腿静脈より2時間ごとに6回静注し,初回注射から16,26,48時間後に血液を採取,血清アルブミン及び血漿フィブリノーゲン濃度を測定した。血清アルブミン値はrhHGF投与後26,48時間目にそれぞれ対照の105,117%に上昇した。血漿フィブリノーゲン値も26,48時間目にそれぞれ対照の141,120%に上昇した。 In vitro実験:ラットからin situでのコラゲナーゼ灌流により肝細胞を単離した。培養肝細胞にrhHGFを添加し,培養上清中アルブミン及びフィブリノーゲン濃度を酵素免疫測定法により測定した。培養上清中アルブミン及びフィブリノーゲン濃度はrhHGFの有無にかかわらず経時的に上昇したが,その程度はrhHGF存在下で有意に大きかった。rhHGF添加量0〜30ng/mlにおいて肝細胞培養上清中のアルブミン濃度はrhHGF 1 ng/mlまで,フィブリノーゲン濃度はrhHGF 10ng/mlまで用量依存性に増加した。アクチノマイシンDの添加により,培養上清中アルブミン及びフィブリノーゲン濃度は低下した。しかし,rhHGFの添加によるアルブミン及びフィブリノーゲン濃度の増加は,アクチノマイシンD添加時にも認められた。アルブミン及びフィブリノーゲンmRNA発現量はrhHGF添加後も非添加に比し,増加しなかった。肝細胞のブロモデオキシウリジン取り込みとアルブミン染色性あるいはフィブリノーゲン染色性とを免疫組織化学的二重染色法にて検討した。アルブミン及びフィブリノーゲン陽性細胞の割合はrhHGF添加後6時間目に非添加群に比し減少した。この時点ではブロモデオキシウリジンの取り込み陽性細胞はみられなかった。rhHGF添加後30時間目にはブロモデオキシウリジン陽性細胞が認められた。このうち12.5%(2/16)のみがアルブミン陽性細胞であった。これに対し,ブロモデオキシウリジン陰性細胞では,77%(214/284)でアルブミン陽性であり,ブロモデオキシウリジン陽性細胞に比し有意にアルブミン陽性率が高かった。一方,フィブリノーゲンの陽性率は,ブロモデオキシウリジン陽性細胞と陰性細胞の間に有意差はみられなかった。 考察:今回のin vivo実験においては,ラットにrhHGFを2時間ごとに6回静注した。この条件では,ブロモデオキシウリジン取り込み陽性肝細胞(S期細胞)及び分裂肝細胞(M期細胞)は,初回注射後26時間目にそれぞれ9.87%と5.92%で,対照群のそれぞれ0%及び0.08%に比して有意に増加し,48時間目には肝DNA量が対照群の127%である。このようなrhHGF投与ラットで,26,48時間目に,血中アルブミン値は対照群の105,117%に,フィブリノーゲン値は141,120%に有意に増加した。静注したrhHGFは肝非実質細胞には取り込まれず肝細胞に取り込まれることから,血中アルブミン及びフィブリノーゲン値の上昇はrhHGFが肝細胞に直接作用し,アルブミン及びフィブリノーゲンの産生ないしは分泌を促進した,あるいはrhHGFにより細胞分裂が起こり蛋白を産生する肝細胞の数が増えた結果であると推定された。 培養肝細胞を用いた実験では,rhHGF添加後24時間目まで培養上清中蛋白濃度が対照に比し増加し,しかも増加分の量も経時的に顕著となった。肝細胞は細胞内に分泌蛋白貯蔵機構を持たず,新たに合成されたアルブミンが細胞外に分泌されるまでの時間は20分以内,フィブリノーゲンにおいても60〜120分とされている。従って,肝細胞からの蛋白の分泌量が増加したのではなく分泌蛋白の合成が亢進した結果と考えられた。またrhHGF添加により添加後3時間目から培養上清中アルブミン及びフィブリノーゲン濃度が増加した。一般にHGF添加後DNA合成が起こるまでには12時間以上が必要であり,今回の実験でもブロモデオキシウリジン取り込み陽性細胞はrhHGF添加後6時間目にみられていない。従って,rhHGFによる蛋白産生促進効果が細胞分裂による細胞数の増加の効果とは考えられなかった。以上から,HGFはDNA合成を促進する一方でアルブミン及びフィブリノーゲン産生も促進すると考えられた。次にDNA合成と蛋白産生が同一の肝細胞で起こるか否かを検討するため,DNA合成の指標であるブロモデオキシウリジン取り込みと細胞質内のアルブミンあるいはフィブリノーゲンに対する免疫組織化学的二重染色を行った。 rhHGF添加後6時間目ではブロモデオキシウリジン陽性細胞はみられなかったがアルブミンあるいはフィブリノーゲン陽性細胞の割合は対照に比べ低下した。HGFを添加した際にブロモデオキシウリジン陽性細胞が出現するのは30時間後であるが,イノシトール三リン酸の産生や細胞内カルシウムイオン動員などの変化は添加後数分以内に始まり,数十分するとc-junやc-fosのmRNA発現もみられる。従って,30時間目でブロモデオキシウリジン陽性となる細胞,すなわちHGFによりDNA合成を開始する細胞は6時間目にすでにDNA合成の準備期にあり,分泌蛋白の産生が抑制されている可能性がある。実際rhHGF添加後30時間目では,アルブミン陽性細胞の割合はブロモデオキシウリジン陰性細胞では陽性細胞に比し高かった。以上より,rhHGFのアルブミン産生促進作用は主として非増殖肝細胞に対するものであると考えられた。しかしフィブリノーゲンに関しては,rhHGF添加後30時間目にはアルブミンと異なりブロモデオキシウリジン陽性,陰性細胞間に染色性の差は認めなかった。フィブリノーゲンはアルブミン同様,肝細胞により合成される蛋白であるが,炎症,外傷等に際して合成が亢進する「急性相蛋白」であり,合成の低下する「非急性相蛋白」であるアルブミンとは逆の合成調節を受けることがある。部分切除後の再生肝においても急性相蛋白のmRNA上昇,非急性相蛋白のmRNA低下がみられる。今回,rhHGFによる肝細胞増殖時にフィブリノーゲンがそれら増殖肝細胞にもみられたことは,増殖時には臓器特有の機能が低下するとする説に合致せず,肝細胞増殖時の分泌蛋白合成は蛋白の種類により異なる調節を受けると推定された。 さらに,培養肝細胞へのrhHGF添加後に,培養上清中のアルブミン及びフィブリノーゲン濃度は有意に上昇しているにもかかわらず,両蛋白のmRNA発現量には増加がみられなかった。また,rhHGFによる培養上清中の両蛋白濃度の増加は,アクチノマイシンDの存在下でも認められた。従って,rhHGFによる両蛋白産生増加は転写後調節によると推定された。 まとめ:HGFは肝細胞のアルブミン及びフィブリノーゲン産生を促進する。その作用機序は転写後の調節である。アルブミン産生促進作用は主として非増殖肝細胞に対するものであるが,フィブリノーゲン産生促進は増殖肝細胞でも同様にみられ,アルブミンの場合とは異なる調節機構の存在が示唆された。 |