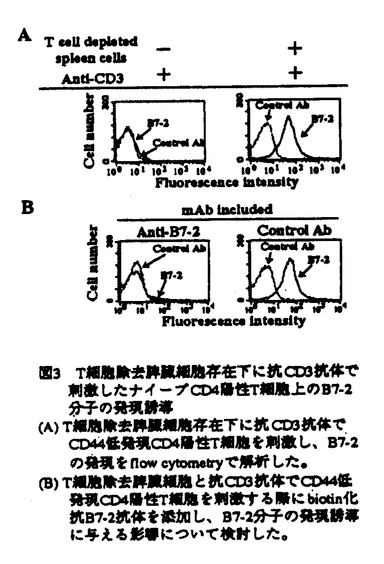
本研究はナイーブCD4陽性T細胞の活性化に重要な役割を果たすB7分子の、CD4陽性T細胞上の発現と機能を明らかにすることを目的として行われたものである。B7-1並びにB7-2分子の発現を、マウス脾臓中のナイーブ/メモリーCD4陽性T細胞亜集団について、細胞表面のCD44並びにCD45RBをナイーブ/メモリー細胞のマーカーとして用いて解析した。さらに、固相化抗CD3抗体でナイーブCD4陽性T細胞を刺激する系を用いて、CD4陽性T細胞上のB7-2分子の機能について、IL-2産生能を指標として解析を試み、下記の結果を得ている。 1.CD4陽性T細胞を用い、ナイーブ/メモリー細胞マーカーとB7分子を蛍光抗体で二重免疫染色し、ナイーブ/メモリーCD4陽性T細胞上のB7分子の発現をフローサイトメーターで解析した結果、B7-2分子はメモリーCD4陽性T細胞上に強く発現しており、ナイーブCD4陽性T細胞上にはほとんど発現していないことが示された。一方、B7-1分子はナイーブ/メモリーCD4陽性T細胞ともに発現が認められなかった。精製したナイーブ/メモリーCD4陽性T細胞を用い、RT-PCR法でmRNAの発現を解析した結果、メモリーCD4陽性T細胞にのみ強いB7-2mRNAの発現が認められ、細胞表面の結果とあわせて、メモリーCD4陽性T細胞上にB7-2分子が発現していることが明らかとなった。 2.B7分子のリガンドであるCD28と免疫グロブリンの融合蛋白を、メモリーCD4陽性T細胞と反応させたところ、結合が認められ、メモリーCD4陽性T細胞上のB7-2分子はCD28分子と結合し得ることが示された。ナイーブCD4陽性T細胞を抗CD3抗体で刺激する際に、メモリーCD4陽性T細胞を共存させた結果、メモリーCD4陽性T細胞非共存時に比べ、有意にIL-2産生が増強することが示された。さらに抗B7-2抗体添加により、メモリーCD4陽性T細胞によるIL-2の産生増強は完全に抑制されることが示され、メモリーCD4陽性T細胞上のB7-2分子が副刺激分子として働き、IL-2産生を増強し得ることが明らかとなった。 3.ナイーブCD4陽性T細胞をT細胞を除去した脾臓細胞存在下で抗CD3抗体刺激し、蛍光抗体で免疫染色し、フローサイトメーターにより解析した結果、CD4陽性T細胞上にB7-2分子の発現が誘導されることが示された。さらに抗B7-2抗体の添加により、CD4陽性T細胞上のB7-2分子の発現誘導は抑制され、ナイーブCD4陽性T細胞上のB7-2分子の発現誘導には脾臓細胞上のB7-2分子による副刺激が必要であることが明らかとなった。ナイーブCD4陽性T細胞を抗CD3抗体とB7-2トランスフェクタントで刺激した結果、B7-2分子の発現が確認された。さらに、ナイーブCD4陽性T細胞を、抗CD3抗体刺激に加え、抗CD28抗体で直接的に副刺激を与えると、B7-2分子が発現誘導されることが示された。 4.ナイーブCD4陽性T細胞を抗CD3抗体とB7-2トランスフェクタントで刺激し、RT-PCR法でCD4陽性T細胞のmRNAの発現を解析した結果、B7-2mRNAの発現が誘導されることが示された。一方、B7-2トランスフェクタント非共存下では、CD4陽性T細胞にB7-2mRNAの発現は誘導されず、抗CD3抗体による刺激とCD28からの副刺激で誘導されるCD4陽性T細胞上のB7-2の発現は、mRNAレベルで誘導されていることが明らかとなった。 5.ナイーブCD4陽性T細胞を抗CD3抗体とB7-2トランスフェクタントで刺激し、経時的にCD4陽性T細胞上のB7-2分子の発現を解析した結果、B7-2分子の発現は刺激後36時間でピークとなり、その後少なくとも2週間後まで発現が持続することが示された。 以上、本論文はマウスCD4陽性T細胞において、ナイーブ/メモリー細胞という活性化の段階の異なる亜集団の解析から、これまで非常に弱く発現していると考えられていたT細胞上のB7-2分子が、活性化の段階の進んだメモリーCD4陽性T細胞で強く発現されていることを証明し、また全く知られていなかった発現誘導のメカニズムを明らかにした。これに加え、これまでT細胞上のB7-2分子は低発現であるため免疫反応を促進する働きはないと報告されていたが、本論文ではメモリーCD4陽性T細胞を精製することにより、T細胞上のB7-2分子が機能的に副刺激分子として働くことを証明し、新たなナイーブ/メモリーT細胞間の相互作用が存在する可能性を示した。このように、本研究は未だほとんど明らかにされていない、CD4陽性T細胞上のB7分子による免疫反応の調節機構の解明に重要な貢献をなすと考えられ、学位の授与に値するものと考えられる。 | 
 g/well)で活性化し、刺激後40時間並びに68時間のIL-2産生量を調べた。この際にパラホルムアルデヒドで固定したCD45RB低発現CD4陽性T細胞を共存させると、非共存下に比べ有意にIL-2の産生量が増加した。このIL-2の産生増強は抗CD3抗体量が30ng/well以上で、刺激後40時間後、68時間後でともに認められた。このCD45RB低発現CD4陽性T細胞によるIL-2の産生上昇は、添加するCD45RB低発現CD4陽性T細胞数に依存し、103cell/cultureでほぼプラトーに達した。
g/well)で活性化し、刺激後40時間並びに68時間のIL-2産生量を調べた。この際にパラホルムアルデヒドで固定したCD45RB低発現CD4陽性T細胞を共存させると、非共存下に比べ有意にIL-2の産生量が増加した。このIL-2の産生増強は抗CD3抗体量が30ng/well以上で、刺激後40時間後、68時間後でともに認められた。このCD45RB低発現CD4陽性T細胞によるIL-2の産生上昇は、添加するCD45RB低発現CD4陽性T細胞数に依存し、103cell/cultureでほぼプラトーに達した。