| 1.はじめに 極端に高いCO2濃度(20-60%CO2)は一般に微細藻類の生育を阻害するストレス要因である。このような極端な高CO2条件下でも生育可能な海産性の単細胞緑藻Chlorococcum littoraleが発見され、その適応機構の解明を目的として様々な研究が行われてきたが、これまでは主に生理・生化学的解析に限られていた。本論文では、分子生物学的手法を用いて極端な高CO2誘導性遺伝子をクローニングし、その遺伝子の発現および酵素タンパク質の誘導条件について検討することを目的とした。さらに、極端な高CO2条件移行時に見られる細胞の微細構造の観察から液胞の形態変化や液胞のマーカー酵素であるV-ATPaseの変化と、これらの極端な高CO2条件への適応の相互関係ついても考察した。 2.極端な高CO2条件誘導性遺伝子のクローニングとその特性 ディファレンシャルスクリーニング法により、20%CO2条件で特異的に発現する2つのcDNAクローンHCR(high-CO2 response)1とHCR2を検出した(表1)。HCR1については、データベースにおいて相同性をもつタンパク質を検索できなかった。一方、HCR2のアミノ酸配列はヒトphagocyteのNADPH oxidaseの大サブユニット(gp91-phox)、酵母の細胞表層ferric reductase(FRE1,FRE2およびFrp1)と部分的にではあるが相同性が見出された。これらのタンパク質は細胞膜を介して細胞外基質に電子を受け渡す役割をもつfravocytochrome familyに属する。また、HCR2と相同性のある領域は、FADおよびNADPH結合に関与すると考えられており(図1)、C.littoraleのHCR2タンパク質も膜上での電子伝達に関与するものと推定した。  表1 High-CO2-response(HCR)cDNAクローンの特徴 表1 High-CO2-response(HCR)cDNAクローンの特徴 図1 HCR2と相同性を示すアミノ酸配列の領域gp91-phox:ヒトphagocyte NADPH oxidaseの大サブユニットFRE1:酵母Saccharomyces cerevisiae FRE2の細胞表層ferric reductase Frp1:酵母Schizosaccharomyces pombeの細胞表層ferric reductase 図1 HCR2と相同性を示すアミノ酸配列の領域gp91-phox:ヒトphagocyte NADPH oxidaseの大サブユニットFRE1:酵母Saccharomyces cerevisiae FRE2の細胞表層ferric reductase Frp1:酵母Schizosaccharomyces pombeの細胞表層ferric reductase HCR2がferric reductaseである可能性が示唆されたことから、C.littoraleにおけるHCR遺伝子発現に及ぼす鉄イオンの影響を調べた。0.036%CO2を含む通常の空気(air条件)で生育した細胞を新鮮培地(10 M FeSO4を含む)に移し、airまたは20%CO2を含む空気(20%CO2条件)を通気して培養を開始した。ノーザンプロットの結果、20%CO2条件に移行した細胞ではそれぞれ2日目と3日目からHCR1および2の発現が認められた(図2)。これに対して、air条件では9日目までそれらの発現が検出されなかった。種々の鉄濃度(0,10,100M FeSO4)存在下で通気をairから20%CO2条件に移行した場合、培地中のFeSO4濃度が高い程HCR1および2遺伝子の発現が抑制された(図3)。これらの結果はHCR遺伝子の発現が高濃度CO2のみならず、培地中の鉄イオン濃度にも影響されることを示す。 M FeSO4を含む)に移し、airまたは20%CO2を含む空気(20%CO2条件)を通気して培養を開始した。ノーザンプロットの結果、20%CO2条件に移行した細胞ではそれぞれ2日目と3日目からHCR1および2の発現が認められた(図2)。これに対して、air条件では9日目までそれらの発現が検出されなかった。種々の鉄濃度(0,10,100M FeSO4)存在下で通気をairから20%CO2条件に移行した場合、培地中のFeSO4濃度が高い程HCR1および2遺伝子の発現が抑制された(図3)。これらの結果はHCR遺伝子の発現が高濃度CO2のみならず、培地中の鉄イオン濃度にも影響されることを示す。  図2 CO2条件を変えたときのHCR-mRNAの発現 図2 CO2条件を変えたときのHCR-mRNAの発現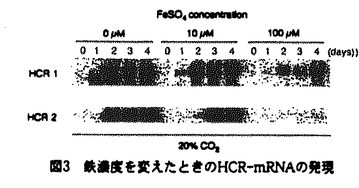 図3 鉄濃度を変えたときのHCR-mRNAの発現 図3 鉄濃度を変えたときのHCR-mRNAの発現 細胞表層ferric reductase活性はair生育細胞では検出できなかったが、20%CO2移行後3日目から増加した(図4)。これはHCR2-mRNAの発現パターン(図2)と同じであった。さらに、HCR2の発現およびferric reductase活性の誘導は、誘導途中のCO2濃度の減少と鉄溶液の添加のいずれによっても抑制された(図5)。この結果は、HCR2のポリクローナル抗体を用いたHCR2タンパク質の誘導パターンと一致したことからも確認された。以上の結果は、HCR2タンパクが細胞膜ferric reductaseである可能性を強く支持すると判断した。  図4 細胞表層ferric reductase活性の誘導経時変化は図2の条件に対応する。 図4 細胞表層ferric reductase活性の誘導経時変化は図2の条件に対応する。 図5 20%CO2・鉄欠乏生育細胞をair条件または鉄添加培地に移行したときのHCR2発現(A)およびferric reductase活性の誘導(B)3.鉄の還元および取り込み活性へおよぼす極端な高CO2および鉄濃度の影響 図5 20%CO2・鉄欠乏生育細胞をair条件または鉄添加培地に移行したときのHCR2発現(A)およびferric reductase活性の誘導(B)3.鉄の還元および取り込み活性へおよぼす極端な高CO2および鉄濃度の影響 細胞表層に誘導される鉄還元活性と鉄代謝との関連を調べるため、ferric reductase活性誘導と細胞への鉄イオンの取り込み速度に対するCO2と鉄イオン濃度の影響を調べた。Air生育細胞(10M FeSO4を含む培地中で培養)をそれぞれ20%CO2/鉄欠乏培地(20%CO2/-Fe)、20%CO2/100M FeSO4添加培地(20%CO2/+Fe)、および通常の空気/鉄欠乏培地(air/-Fe)条件に移し各活性を測定した。鉄の取り込み活性は、ferric reductase活性の影響を除外する目的から、反応液中にアスコルビン酸を添加して59Fe(III)を59Fe(II)に還元した条件で細胞内へ取り込まれた放射活性から求めた。 59Feの直線的な取り込みは、20%CO2/-Fe細胞でのみ観察された。また、取り込み活性がFe(II)に特異的なキレート剤での添加によって完全に抑制されたことから、鉄の取り込みはFe(III)ではなく、Fe(II)イオンの形で起こっていることが示された。鉄取り込みの初期速度に対するFe(II)濃度の影響を調べると、いずれの条件でも一定のFe(II)濃度で飽和に達し、曲線はMichaelis-Mentenの式を示した。概算されたKmは鉄欠乏条件(20%CO2/-Feとair/-Fe)で鉄添加生育条件(20%CO2/+Fe)よりも1/30と低く、鉄への親和性が高いことを示していた(表2)。また、20%CO2条件では鉄の有無に依存せず、air条件よりも5倍高いVmax値を示した。このことから、極端な高CO2/鉄欠乏生育細胞では、鉄への親和性の増大および鉄の蓄積量の増大の双方が起きてる事が明らかとなった。  表2 Fe(II)取り込み初速度の鉄濃度に対するKmとVmax値r2はnonlinear carve fittingの相関係数を示す。 表2 Fe(II)取り込み初速度の鉄濃度に対するKmとVmax値r2はnonlinear carve fittingの相関係数を示す。 20%CO2/-Fe条件では、2日目にFe(II)取り込みの上昇が見られたが、air/-Fe条件および20%CO2/+Fe条件では顕著な変化は見られなかった(図6A)。細胞表層ferric reductaseは20%CO2/-Fe条件に移行した場合にのみ2日目に活性の上昇そ示した(図6B)。以上の結果は、20%CO2および鉄欠乏条件の双方が与えられた場合にのみ鉄の取り込みとferric reductase活性の誘導が起こることを示す。  図6 Fe(II)取り込み初速度(A)とferric reductase活性(B)誘導の経時変化 図6 Fe(II)取り込み初速度(A)とferric reductase活性(B)誘導の経時変化 光合成電子伝達系の阻害剤(DCMU)の添加、および光の有無はFe(II)の取り込みに影響を与えなかったが、膜結合性ATPaseの阻害剤(DCCD)や脱共役剤(CCCP)では鉄取り込み活性の顕著な低下が認められた(表3)。この結果は、細胞への鉄取り込みがATPaseを介したエネルギー要求性の機構である事を示唆する。また、ferric reductase活性がDCMUやDCCDには影響されず、CCCPにより阻害された(表3)事から、膜内外のpH差が本酵素の活性化に影響することを示す。  表3 細胞表層ferric reductase活性およびFe(II)取り込み初速度への阻害剤の影響・暗条件で測定を行った。・コントロールに対する活性の割合を示す。4.極端な高CO2条件におけるChlorococcum littoraleの液胞数と液胞型ATPase量の増加 表3 細胞表層ferric reductase活性およびFe(II)取り込み初速度への阻害剤の影響・暗条件で測定を行った。・コントロールに対する活性の割合を示す。4.極端な高CO2条件におけるChlorococcum littoraleの液胞数と液胞型ATPase量の増加 極端な高CO2条件で生育したC.littorale細胞では、液胞の形態変化が特に40%CO2条件下で起こることを見出した。Airから40%CO2条件へ移行した時、統計的に計測した液胞数と断面積はいずれも増加することを明らかとした(図7)。  図7 40%CO2およびair条件における細胞の生育(A)、液胞の数(B)と面積(C)の経時変化 図7 40%CO2およびair条件における細胞の生育(A)、液胞の数(B)と面積(C)の経時変化 さらに、高等植物ヤエナリの液胞型ATPaseのサブユニットB抗体により、C.littoraleの同タンパク質の検出を試みた結果、ヤエナリとほぼ同じ分子量に交差反応するバンドが検出された(図8A)。この液胞型ATPaseのバンドの経時変化は、air条件で生育した細胞では8日間ほとんど変化が見られなかったのに対し、40%CO2条件に移行した細胞では約2倍の増加が見られた(図8B)。また、airと40%CO2条件で8日間生育した細胞の液胞膜画分の液胞型ATpase活性(硝酸塩感受性のATPase活性)をそれぞれ測定したところ、40%CO2生育細胞の液胞画分にはair生育細胞のそれに比べ約2倍の活性が検出された。これらの結果は、C.littoraleのATPaseを付随した液胞の増大が極端な高CO2条件に適応する間に起こっていることを示している。  図8 ヤエナリ抗体によるV-ATPase subunit Bのイムノブロット(A)およびバンドの相対量(B)の経時変化各CO2条件で生育した細胞の全タンパク質10gをアプライした。MBはヤエナリ液胞のタンパクを示す。5.おわりに 図8 ヤエナリ抗体によるV-ATPase subunit Bのイムノブロット(A)およびバンドの相対量(B)の経時変化各CO2条件で生育した細胞の全タンパク質10gをアプライした。MBはヤエナリ液胞のタンパクを示す。5.おわりに C.littoraleから極端な高CO2と鉄欠乏条件下でその発現が促進される2つのHCR遺伝子(HCR1と2)をクローニングした。HCR2タンパク質は、(1)酵母FRE1とのアミノ酸配列の相同性。(2)遺伝子発現と酵素活性誘導のパターンの一致から、細胞表層局在のferric reductaseであると予想した。また、HCR2の発現とともに鉄取り込みとferric reductase活性の誘導も同様に見られた事から、HCR2およびferric reductase、鉄の取り込みには強い相関があるものと判断した。極端な高CO2条件では光合成活性や生育速度の増大が伴うことから、HCR遺伝子および酵素活性の誘導は、必要とされる鉄含有酵素タンパクの相対的な量の不足に起因する可能性が考えられる。極端な高CO2条件においては細胞の酸性化が引き起こされると考えられるが、それを抑制し細胞内pHを調節するための機構の一つとして、細胞における液胞および液胞型ATPaseの増大がもたらされると推定した。 本論文で示された極端な高CO2濃度により調節される微細藻類の鉄濃縮機構の誘導や液胞の発達は新たな知見であり、微細藻細胞の有する適応機構の多様性を示唆する。本研究は、極端な高濃度のCO2条件が微細藻類においてストレスとなる原因やそれらストレスへの適応機構の解明に有用な知見を提供するものと思われる。 |