コリネ型細菌はグルタミン酸をはじめとするアミノ酸発酵に広く用いられている。コリネ型細菌のグルタミン酸生成機構は大変興味深く、通常の生育条件下ではグルタミン酸を全く生成しないが、生育必須因子であるビオチンが欠乏した条件下では生育は抑制されるが著量のグルタミン酸を生成する。また、ビオチンが過剰に存在する条件下でもある種の界面活性剤やペニシリンを培地に添加すると、グルタミン酸を著量生成する。さらに、リン脂質の構造に影響を及ぼすと考えられるオレイン酸やグリセロールの要求性変異株も、過剰のビオチン存在下でグルタミン酸を生成するようになる。アミノ酸発酵の研究が盛んに行われた1960年代は、これらのグルタミン酸生成を引き起こす因子が全て細胞表層の構造に影響を及ぼすと予想された事から、これらの因子により細胞質膜の透過性が上昇し、グルタミン酸が細胞質膜の脂質二重層から漏出してくると考えられた。実際にグルタミン酸生成条件下では、リン脂質や脂肪酸の組成が変化するという知見も得られていた。漏出説の根拠となるもう一つの知見は、コリネ型細菌には2-oxoglutarate dehydrogenase complex(ODHC)活性が無いか極めて微弱であるというものであった。ODHCはグルタミン酸の前駆体である2-オキソグルタール酸をスクシニル-CoAに変換するTCA回路の酵素である。つまり、コリネ型細菌ではもともと代謝フラックスがグルタミン酸生成に傾いていることに加え、上記の因子の作用で細胞質膜の透過性が向上することにより、細胞内のグルタミン酸濃度が低濃度に保たれ、グルタミン酸生成が抑制されずグルタミン酸が細胞外に著量蓄積してくると考えられたのである。 しかし、その後の研究により漏出説を否定する知見が多く得られてきた。まず、コリネ型細菌でもODHC活性を安定に測定できる系が確立され、代謝フラックスがグルタミン酸生成に傾いているわけでは無いことが示された。また、膜透過機構についても研究が進み、ビオチン制限条件下においてグルタミン酸以外の物質の膜透過性は変化しないことが示され、さらにグルタミン酸の取り込み系とは別の排出系が存在することが生化学的および遺伝学的解析から明らかになった。 近年、コリネ型細菌のグルタミン酸生成においてODHCの活性とグルタミン酸生成との関連に着目した研究がなされ、ビオチン制限、ペニシリン添加、界面活性剤添加によるグルタミン酸生成条件下においてODHC活性が1/3程度に低下している事が判明した。これらの知見からコリネ型細菌のグルタミン酸生成において、ODHC活性の低下が重要な意味を持っていると考えられた。 こうした背景のもとで,コリネ型細菌に特有なグルタミン酸生成は一体どのような機構で引き起こされているのかが改めて疑問となった。本研究はこの疑問を解明するために行った。 まず、界面活性剤によるグルタミン酸生成機構に着目して研究を開始した。コリネ型細菌は充分量のビオチン存在下ではグルタミン酸を生成せず菌体形成のみがおこる。しかし、適量のTween40を添加すると1〜2回分裂した後に生育は停止し、グルタミン酸の過剰生成が誘導される。そこでTween40によるグルタミン酸生成に関与する遺伝子の取得を試みた。 コリネ型細菌野生株Corynebacterium glutamicum(Brevibacterium lactofermentum)ATCC13869株から得られたTween40感受性変異株AJ11060株の、Tween40感受性を相補する遺伝子を野生株の染色体遺伝子ライブラリーから取得した。この遺伝子断片(2855bp)の塩基配列を決定したところ、543アミノ酸からなるタンパク質をコードすると予想されるORFが一つ検出された。推定されたアミノ酸配列を基に部位特異的抗体を作製し、ウエスタンブロッティングによりこの遺伝子の発現を確認した。この遺伝子をdtsR遺伝子(Rescurer gene of Detergent sensitivity)と命名した。 DtsRの機能を推測すべく塩基配列から推定されたアミノ酸配列を基に相同性の高い既知タンパク質を検索した。相同性の高いものがいくつか検出されたが、興味深いことにそれらは全てビオチン酵素として知られているものであった。高い相同性が見られたのはラットおよびヒト由来のPropionyl-CoA carboxylase(PCC)の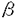 サブユニット(PccB)であった。いずれも全長にわたり約50%のアミノ酸が一致していた。特に機能領域と考えられているカルボキシルビオチン結合領域やCoA結合領域は高く保存されていた。 サブユニット(PccB)であった。いずれも全長にわたり約50%のアミノ酸が一致していた。特に機能領域と考えられているカルボキシルビオチン結合領域やCoA結合領域は高く保存されていた。 一方、dtsR遺伝子取得の報告をしたのとほぼ同時期に、PuhlerらはC.glutamicumのビオチン結合タンパク質としてAccBC(acyl-CoA carboxylase subunit B and C)の取得を報告した。AccBCはPCCの サブユニット(PccA)と高い相同性を示す。PccAとPccBとは複合体を形成していることが知られており、AccBCとDtsRも複合体を形成している可能性が高いと考えられた。 サブユニット(PccA)と高い相同性を示す。PccAとPccBとは複合体を形成していることが知られており、AccBCとDtsRも複合体を形成している可能性が高いと考えられた。 dtsR遺伝子とグルタミン酸生成との関係を調べるに当り、dtsR遺伝子増幅株のグルタミン酸生産性をビオチン制限法、界面活性剤添加法およびペニシリン添加法において調べた。その結果、界面活性剤添加法のみならずビオチン制限法やペニシリン添加法においてもグルタミン酸生成の著しい低下が観察された。この結果からDtsRがコリネ型細菌のグルタミン酸生成機構において何らかの中心的な役割を果たしていると考えられた。また、dtsR増幅がグルタミン酸生成に抑制的に働くことから逆にdtsR遺伝子を破壊することによりグルタミン酸生成が促進される可能性が示唆された。 そこで、C.glutamicum ATCC13869株よりdtsR破壊株の作製を試みた。検討当初は、dtsR破壊株を取得することが出来なかったが、DtsRがビオチン酵素であり、脂肪酸合成に関与する可能性が考えられた。そこで、選択培地にオレイン酸またはオレイン酸エステルを添加することによりdts破壊株の取得に成功した。 次に、dtsR破壊株のグルタミン酸生成を調べた。dtsR破壊株の生育速度は野生株と比較し低下していたが、本培養条件では野生株はグルタミン酸を生成しないのに対し、dtsR破壊株は著量のグルタミン酸を生成した。このグルタミン酸生成量は野生株のビオチン制限条件とほぼ同程度であった。この結果から、DtsRはコリネ型細菌のグルタミン酸生成に重要な影響をおよぼすことが明らかとなった。上述の通りdtsR破壊株は脂肪酸要求性になることからコリネ型細菌の脂肪酸合成に必須な酵素をコードしていると考えられるが、同時にグルタミン酸生成も引き起こしたことから、コリネ型細菌において脂肪酸合成系とグルタミン酸合成系とがDtsRを介して何らかの関連を持っていることが考えられた。従来、膜構造の変化とグルタミン酸生成を切り離して考えることが出来なかったのはこの為だと思われる。 dtsR破壊によりグルタミン酸を生成するようになったことから、dtsR破壊株においては代謝フラックスがグルタミン酸生成に向かっていると考えられた。そこでdtsR破壊株においてODHC活性が影響を受けているかどうかに興味が持たれた。dtsR破壊株のODHC活性がどのように変化しているかを調べたところ、野生株に比較し比活性が約3分の1に低下していた。 dtsR遺伝子は界面活性剤の機能に着目して取得した遺伝子であり、DtsRはAccBCとビオチン酵素複合体を形成していると考えられた。さらに、dtsR破壊株がグルタミン酸を生成することが明らかとなり、ビオチン制限や界面活性剤添加のターゲットはDtsRを含むビオチン酵素複合体であり、DtsRを含むビオチン酵素複合体の活性が低下することによりグルタミン酸生成を引き起こしていると考えられる(図1)。また、オレイン酸要求株によるグルタミン酸生成もこのビオチン酵素複合体の欠失によるものと考えれば理解できる。以上の結果から、DtsRはコリネ型細菌に特有のグルタミン酸生成の誘導に関与していると推測される。グルタミン酸生成条件下におけるDtsRの機能低下がどのようなメカニズムでODHC活性の低下を引き起こすのかを明らかにすることにより、コリネ型細菌のグルタミン酸生成機構の全貌を明らかにすることが出来ると思われる。  図1 コリネ型細菌のグルタミン酸過剰生成機構 図1 コリネ型細菌のグルタミン酸過剰生成機構 |