本研究は、臓器移植における急性拒絶反応に重要な役割を演じるヒト主要組織適合抗原(HLA)クラスI分子を解析する手段として、二種類のHLAクラスIトランスジェニックマウス(C3H.B51およびC3H.B35)の有用性を解析したものであり、下記の結果を得ている。 1.C3H.B51は、C3H.B35の皮膚を平均11.4日で拒絶した。これは、アロの皮膚の拒絶までの期間とほぼ同等であり、2種類のHLAクラスIトランスジェニックマウスの間で、互いのHLAクラスI分子を非自己として認識することが示された。一方C3H.B51は、C3H.B35の心を平均22.8日で拒絶した。皮膚移植での結果と同様に、2種類のHLAクラスIトランスジェニックマウス間で、互いのHLAクラスI分子をアロ抗原として認識することが示された。 2.免疫組織染色標本では、C3H.B51に移植したC3H.B35の心に著明なCD3陽性CD8陽性細胞の浸潤がみられ、細胞傷害性T細胞が拒絶反応に重要であることが示された。 3.C3H.B51の脾細胞より得られたエフェクター細胞は、C3H.B35リンパ芽球および、L-B*3501細胞に対する細胞傷害性を有していた。このエフェクター細胞が、HLA-B*3501分子自体を認識するのか、あるいはマウスMHC(H-2)分子に提示されたHLA-B*3501分子ペプチドを認識するかを確認するために、H-2分子を発現しないC1R-B*3501細胞をcold targetとして用いたところ、エフェクター細胞のC3H.B35リンパ芽球に対する細胞傷害性は抑制された。これらの結果より、C3H.B51より得られたエフェクター細胞は、H-2分子に提示されたHLA-B*3501分子ペプチドを認識するのではなく、HLA-B*3501分子のpolymorphic regionを直接認識する、つまりアロMHCクラスI抗原として認識することが示された。 4.上記で得られたエフェクター細胞を抗Thy-1抗体と補体で処理することによりT細胞を除去したところ、HLA-B*3501に特異的な細胞傷害性は見られなくなった。またこのエフェクター細胞を抗CD8抗体と補体で処理することによりCD8陽性細胞を除去した場合も細胞傷害性は見られなくなった。一方、エフェクター細胞を抗CD4抗体と補体で処理する事によりCD4陽性細胞を除去しても細胞傷害性は保たれていた。以上の結果より、この細胞傷害性細胞はCD8陽性細胞であることが確認された。 5.C3H.B51マウスのの胸腺内にHLA-*3501分子由来のペプチド(B*3501/101-125)を投与し、48時間後にC3H.B35の心を移植したところ移植組織片の生着期間が延長した。この結果より、アロHLAクラスIペプチドを胸腺内に投与することにより移植組織片の生着期間が延長しうることが示された。 以上、本論文はHLAクラスIトランスジェニックマウスが、移植におけるHLAクラスI分子の解析を行うために有用であることを示し、また、ドナー由来のHLAクラスI分子由来のペプチド投与による寛容誘導の可能性を示唆した。本研究は、移植免疫におけるHLAクラスI分子の役割の解明に重要な貢献をなすと考えられ、学位の授与に値するものと考えられる。 | 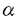 1および
1および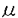 g/マウスのpepC.(B*3501/101〜125)を投与したところ5例すべてが60日以上の移植組織片の生着がみられた。また、pepB(B*3501/81-104)の投与によっても長期生着例がみられた。しかし、他のペプチド投与による効果は明らかではなかった。以上の結果より、アロHLAクラスIペプチドを胸腺内に投与することにより移植組織片の生着延長効果を期待しうることが示唆された。
g/マウスのpepC.(B*3501/101〜125)を投与したところ5例すべてが60日以上の移植組織片の生着がみられた。また、pepB(B*3501/81-104)の投与によっても長期生着例がみられた。しかし、他のペプチド投与による効果は明らかではなかった。以上の結果より、アロHLAクラスIペプチドを胸腺内に投与することにより移植組織片の生着延長効果を期待しうることが示唆された。