| 1.序論 生物における学習がどのように成立し、記憶としてどのように蓄えられるのか、これまでに様々な動物において、種々の切り口から数多くの研究がなされてきた。軟体動物は、その神経系が比較的単純であるにもかかわらず、数多くの学習を行う能力を有することから、学習・記憶の研究に非常によく利用されている。本研究では非常に発達した嗅覚神経系を有する陸棲の軟体動物であるコウラナメクジの行動実験を通して学習・記憶のメカニズムを考察した。まず匂いと味による古典的条件付けを行い、多様な学習の組み合わせとその結果見られる行動変化から、記憶間の関係性およびその時間変化を調べた。また、代謝活性マーカーを使用することで、嗅覚中枢である前脳葉において、匂い学習に関係する細胞群をとらえることに成功した。さらに、神経可塑性と関連の深い神経修飾物質であるセロトニンとFMRFアミドの免疫抗体を用いて組織学的に嗅覚神経系の神経構造を観察し、それらの物質が前脳葉に特徴的な分布をしていることを明らかにした。これらの結果をもとに、行動-神経回路網-細胞レベルからコウラナメクジの匂い学習のメカニズムについて考察する。 2.冷却による逆行性健忘を用いたパブロフ条件付けの刺激間連合の研究 これまでの研究からコウラナメクジは古典的条件付けにより2つの刺激間の関係性を学習する能力を有すること、さらに、条件刺激を与えて記憶を活性化した直後に冷却を施すことで蓄えられた記憶に逆行性健忘が起きることなどが分かっている。そこで、ナメクジに匂い-味忌避条件付け学習を行った後に、冷却による逆行性健忘を起こし、その健忘の起き方から記憶間の関係性を調べた。 条件付けには条件刺激として2つの匂い(ニンジン:CS1,キュウリ:CS2)と無条件刺激として苦味の物質である硫酸キニジン(US)を用い、独立1次条件付け(CS1+US,CS2+US)、順次型2次条件付け(CS1+US,CS2+CS1)、同時型2次条件付け(CS1+US,[CS1,CS2])、および感性的予備条件付け(CS2+CS1,CS1+US)の4通りの条件付け学習を図1に示す時間スケジュールで行った。いずれの条件付けでもCS1、CS2ともに忌避反応を示したが、条件付け後に、条件刺激に用いた各匂いを与えてから冷却を行い逆行性健忘を引き起こすと、健忘の起き方が条件付けの種類によって異なっていた(表1)。すなわち学習の結果として同じ忌避反応を示す記憶でも、コウラナメクジの脳内における記憶の蓄えられ方は異なった関係性を持っていることが明らかになった。 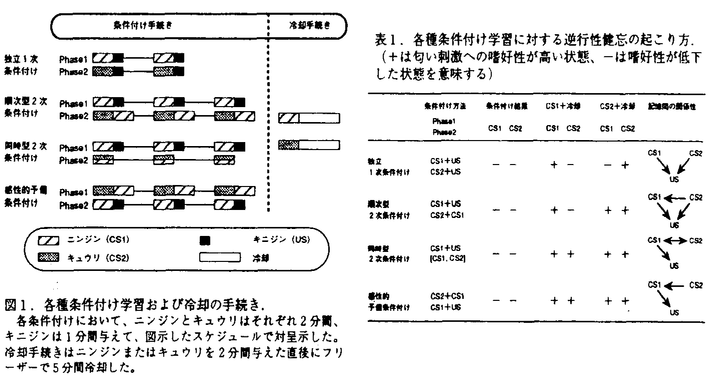 図表図1.各種条件付け学習および冷却の手続き. (+は匂いの刺激への嗜好性が高い状態、-は嗜好性が低下した状態を意味する) / 表1.各種条件付け学習に対する逆行性健忘の起こり方. 各条件付けにおいて、ニンジンとキュウリはそれぞれ2分間、キニジンは1分間与えて、図示したスケジュールで対呈示した。冷却手続きはニンジンまたはキュウリを2分間与えた直後にフリーザーで5分間冷却した。3.再学習による記憶の状態変化 図表図1.各種条件付け学習および冷却の手続き. (+は匂いの刺激への嗜好性が高い状態、-は嗜好性が低下した状態を意味する) / 表1.各種条件付け学習に対する逆行性健忘の起こり方. 各条件付けにおいて、ニンジンとキュウリはそれぞれ2分間、キニジンは1分間与えて、図示したスケジュールで対呈示した。冷却手続きはニンジンまたはキュウリを2分間与えた直後にフリーザーで5分間冷却した。3.再学習による記憶の状態変化 コウラナメクジにおける行動学的研究から、冷却により逆行性健忘が引き起こせるのは、学習後3日程度までの新しい記憶に限られることが分かっている。そこで順次型2次条件付け手続きを利用し、図2に示すような実験スケジュールに従って2つのフェーズ間の間隔(日数)を変えることで、古い記憶(Phase1)と新しい記憶(Phase2)の時間変化を調べた。その結果、本来なら逆行性健忘が起こらない1週間を経過した古い記憶でも、その古い記憶を用いた新しい記憶が形成された時には、古い記憶にも健忘が起きることが分かった(図3)。このことは新しい記憶が形成されるときに古い記憶も含めて同時に形成し直され、全体が新しい状態(冷却によって逆行性健忘が引き起こされる状態)として記憶し直されることを意味している。  図表図2.順次型2次条件付けにおけるPhase間隔を変えた実験スケジュール. 学習はPhase1としてCS1+USを2回、Phase2としてCS2+CS1を2回行い、Phase間の間隔を1、3、6、7日とした4グループを作った。 / 図3.新しい記憶(SOC)の古い記憶(FOC)への影響(CS1+F冷却手続き). 順次型2次条件付け学習のPhase1(1次条件付け:FOC)とPhase2(2次条件付け:SOC)との間隔を1、3、6、7日のグループを作り、全グループの条件付け終了後、CS1+Fの冷却手続きにより逆行性健忘を引き起こした。匂い嗜好性テストの結果、Phase1の学習後6、7日を経過して古い記憶となっているはずのCS1の記憶に健忘が起こり、CS1に対する嗜好性が上昇している。右の棒グラフは2次条件付けグループとControlグループの匂い嗜好性を示すもの。(*:t-test p<0.01)4.嗅覚学習に関係した前脳葉インターニューロン群の機能構造 図表図2.順次型2次条件付けにおけるPhase間隔を変えた実験スケジュール. 学習はPhase1としてCS1+USを2回、Phase2としてCS2+CS1を2回行い、Phase間の間隔を1、3、6、7日とした4グループを作った。 / 図3.新しい記憶(SOC)の古い記憶(FOC)への影響(CS1+F冷却手続き). 順次型2次条件付け学習のPhase1(1次条件付け:FOC)とPhase2(2次条件付け:SOC)との間隔を1、3、6、7日のグループを作り、全グループの条件付け終了後、CS1+Fの冷却手続きにより逆行性健忘を引き起こした。匂い嗜好性テストの結果、Phase1の学習後6、7日を経過して古い記憶となっているはずのCS1の記憶に健忘が起こり、CS1に対する嗜好性が上昇している。右の棒グラフは2次条件付けグループとControlグループの匂い嗜好性を示すもの。(*:t-test p<0.01)4.嗅覚学習に関係した前脳葉インターニューロン群の機能構造 上記の学習行動の神経メカニズムを調べるために、ナメクジが解放血管系であることを利用して、匂い-味条件付け学習を行った直後に代謝活性マーカーとして蛍光性の色素ルシファーイエローをナメクジの腹腔内に注入し、嗅覚学習に関係した代謝活性の高い神経細胞内へ色素が取り込まれる様子を観察した(図4)。その結果、忌避学習を行った個体では未処理やコントロールの個体に比べて、嗅覚中枢である前脳葉で非常に多くのインターニューロンがラベルされていた。さらに、忌避学習個体の前脳葉で色素の取り込みを示していた神経細胞は、背腹軸に平行方向に帯状のクラスターを形成していた(図5)。これらの細胞群は記憶の形成に際してその代謝活性が大きな影響を受けた部分と考えられ、その帯は前脳葉で見られる電位振動の位相がそろっている方向と一致していた。コウラナメクジの前脳葉における電位振動パターンは嗅覚学習をこより変化することが分かっており、背腹軸に平行な細胞群は匂いに関する学習・記憶において重要な役割を担う1つの機能単位であると考えられる。  図4.1次条件付け学習時における前脳葉細胞内への色素取り込み実験.aに示すスケジュールにしたがって、条件付け([CaQ])、対照([Ca-Q])、および未処理(Naive)の3グループについて実験を行った。条件付け終了20分後にルシファーイエロー(LY)溶液を体腔に注入し、その1時間後に解剖してラベルされている前脳葉の内在性神経細胞を計数した。bのグラフは左右の前脳葉のうち、ラベルされている細胞数が多い方の数値をその個体のラベル細胞数として取ったときの、前脳葉当たりの平均数を示す。 図4.1次条件付け学習時における前脳葉細胞内への色素取り込み実験.aに示すスケジュールにしたがって、条件付け([CaQ])、対照([Ca-Q])、および未処理(Naive)の3グループについて実験を行った。条件付け終了20分後にルシファーイエロー(LY)溶液を体腔に注入し、その1時間後に解剖してラベルされている前脳葉の内在性神経細胞を計数した。bのグラフは左右の前脳葉のうち、ラベルされている細胞数が多い方の数値をその個体のラベル細胞数として取ったときの、前脳葉当たりの平均数を示す。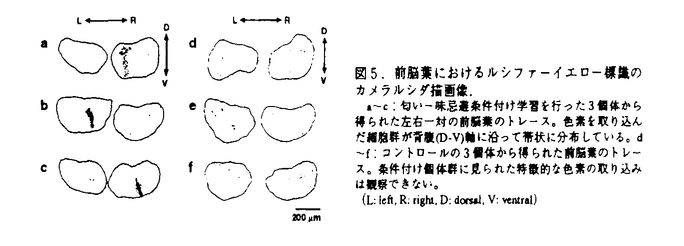 図5.前脳葉におけるルシファーイエロー標識のカメラルシダ描画像.a〜c:匂い-味忌避条件付け学習を行った3個体から得られた左右一対の前脳葉のトレース。色素を取り込んだ細胞群が背腹(D-V)軸に沿って帯状に分布している。d〜f:コントロールの3個体から得られた前脳葉のトレース。条件付け個体群に見られた特徴的な色素の取り込みは観察できない。(L:left,R:right,D:dorsal,V:ventral)5.コウラナメクジ嗅覚神経系における免疫組織学的研究 図5.前脳葉におけるルシファーイエロー標識のカメラルシダ描画像.a〜c:匂い-味忌避条件付け学習を行った3個体から得られた左右一対の前脳葉のトレース。色素を取り込んだ細胞群が背腹(D-V)軸に沿って帯状に分布している。d〜f:コントロールの3個体から得られた前脳葉のトレース。条件付け個体群に見られた特徴的な色素の取り込みは観察できない。(L:left,R:right,D:dorsal,V:ventral)5.コウラナメクジ嗅覚神経系における免疫組織学的研究 コウラナメクジの前脳葉は匂い学習において重要な組織であることが分かったので、嗅覚神経系の中で前脳葉および嗅覚刺激の入力部位である触角において、免疫組織学的手法を用い、神経修飾物質候補であるFMRFアミドとセロトニンについての分布を観察した。 コウラナメクジの嗅覚中枢である前脳葉は、インターニューロンの細胞体から成るCell mass(CM)、触角神経節からの投射部位(嗅覚情報の入力部位)であるTerminal mass(TM)および前脳葉と中脳葉の接合部位であるIntemal mass(IM)の3つの部域に構造上分かれている。CMが細胞体から成るのに対し、TMとIMはCMからの神経線維以外にも池の神経節から投射する多くの線維が絡み合うニューロパイルである。この前脳葉において、セロトニン様免疫活性を示す線維が前脳葉ニューロパイル部域全体に広く分布していたが、その中でも特にTMではIMよりも高密度に分布しておりvaricosity構造も持っていた。一方、FMRFアミド様免疫活性を示したのは2種類の線維で、1つはCM内で背腹軸と平行方向に分枝している線維と、もう1つはIMと中脳葉との接合部でvaricosity構造を示す線維が観察された。いずれの線維も細胞体は前脳葉内には観察されず、別の神経節から投射しているものであった。これらの形態的特徴から、セロトニン様関連物質は触角から前脳葉への入力に対して修飾している可能性がある。またCM内で背腹軸方向に分枝するFMRFアミド様線維の分布は、記憶に関係した機能単位の方向と一致しており、それらの細胞を修飾している可能性がある。 次に、嗅覚刺激の入力部位にあたる触角神経節にはFMRFアミド様免疫陽性の一次感覚神経細胞が見つかったが、嗅覚中枢である前脳葉への投射は観察されなかった。また、セロトニン様陽性細胞は触角神経節には見られず、中枢側から投射している線維のみがニューロパイル内に観察された。 6.結論 行動実験の結果から、(1)記憶は時間とともに変化しており、刺激間の関係は呈示される順序によって異なった連合を形作ること、さらに、(2)古い記憶はそのまま保存され続けるのではなく、新しい記憶が形成されるときにそれらと一緒に新しい状態へ形成し直されることが分かった。これらの結果は、ナメクジが多様な環境の変化に適応して生きていく際に、経験に基づいて入力を解釈し、新しい記憶を作っていく過程を反映したものと考えられる。 また嗅覚中枢の前脳葉を中心とした組織学的な観察結果から、図6に示すような嗅覚学習に関係する神経ネットワークモデルが考えられる。コウラナメクジの前脳葉で見られる電位振動パターンは匂い刺激や学習の前後で変調されることが見つかっており、セロトニンやFMRFアミドが神経修飾物質として動機付けや強化の過程に介在することで、それらの物質が前脳葉の電位振動のパターンを変化させていると考えられる。このような神経ネットワークのダイナミックな変化がコウラナメクジの柔軟な記憶を形成しているものと考えられる。  図6.コウラナメクジの匂い-味条件付け学習モデル.触角で受容された匂い刺激(CS)の情報は前脳葉で処理され、その結果に基づき行動が発現していると考えられる。味(US)に関する情報は行動の発現を左右する動機付けの過程に作用し、そこから情動系の情報が前脳葉や触角神経節に入力する。セロトニンやFMRFアミド関連物質は動機付け・強化過程に含まれていると考えられ、それらの神経修飾質が前脳葉や触角神経節へのフィードバック経路に介在することで、シナプスの可塑性を含む全体のダイナミクスの変化が生じると考えられる。 図6.コウラナメクジの匂い-味条件付け学習モデル.触角で受容された匂い刺激(CS)の情報は前脳葉で処理され、その結果に基づき行動が発現していると考えられる。味(US)に関する情報は行動の発現を左右する動機付けの過程に作用し、そこから情動系の情報が前脳葉や触角神経節に入力する。セロトニンやFMRFアミド関連物質は動機付け・強化過程に含まれていると考えられ、それらの神経修飾質が前脳葉や触角神経節へのフィードバック経路に介在することで、シナプスの可塑性を含む全体のダイナミクスの変化が生じると考えられる。 コウラナメクジ前脳葉で見られる電位振動パターンの変化がどのように行動の変化に反映されているかはまだ詳しく分かっていないが、匂いの記憶に関係する機能単位の発見は生理学的なアプローチの可能性を提起しており、コウラナメクジの嗅覚学習は行動レベルからの研究と細胞レベルからの研究とを結びつけるのに非常に有用な実験系であると思われる。今後は、様々な条件付け学習において前脳葉の機能単位を中心とした生理学的アプローチを行うことで、神経ダイナミクスの変化と記憶コーディングとの関係を明確にできるものと考えられる。 |