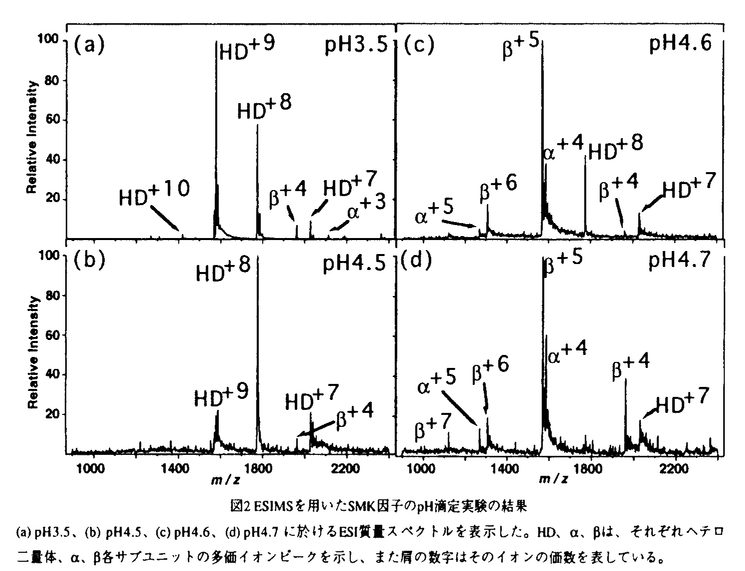
| 審査要旨 | | 酵母などのある種の株は,キラー因子とよばれる蛋白質を分泌し,競合する他の株などを死滅させる。味噌麹中より見い出された耐塩性酵母Pichia farinosa KK1株が産生するSMKキラー因子は,一本鎖の前駆体蛋白質が翻訳後修飾を受け, と と のサブユニットから成る分子量約14,000の成熟型蛋白質として分泌されて,キラー活性を示す。その作用機序に関しては,感受性株のアデノシントリホスファターゼとの関連や,細胞膜に対する非特異的な破壊作用などが指摘されている。 のサブユニットから成る分子量約14,000の成熟型蛋白質として分泌されて,キラー活性を示す。その作用機序に関しては,感受性株のアデノシントリホスファターゼとの関連や,細胞膜に対する非特異的な破壊作用などが指摘されている。 酸性下で構造的に安定なSMK因子の作用機序を三次元構造の知見に基づいて明らかにすることは,有用微生物の選択的な利用のみならず,細胞生物学の諸相の理解にも重要である。本論文の研究では,X線結晶構造解析によってSMK因子の三次元構造を解明し,さらに,質量分析法等を併用してその構造安定性などを解析している。 SMKキラー因子の三次元構造 研究では,まず,硫酸アンモニウム(AS)溶液により析出させた正方晶系に分類されるAS析出結晶について,多重重原子同型置換法のX線結晶構造解析によって,成熟型SMK因子の三次元構造(挿入図に示す)を分解能1.8Åで解明した。構造は,と両サブユニットが協同して折り畳んだ,単一のドメインからなる楕円体型をとり,5本鎖の逆平行シートの片面に2本のヘリックスが並んで配置する/サンドイッチの折りたたみに二次構造的には分類される。SMK因子の三次元構造は担子菌Ustilago maydisが産するKP4キラー因子との類似性を有し,両者の間には進化的および機能的に密接な関連性があることも見出している。  図表 図表 SMK因子の前駆体には,とのサブユニットの間に,約60残基からなる ペプチドが介在する。サブユニットのC末端とサブユニットのN末端は,成熟型分子の上部に近接していることから,前駆体分子の上部を覆い隠すように独立したドメインとして存在しているペプチドが切り出されて露出する分子の上部表面が活性の発現に重要であると考察している。 ペプチドが介在する。サブユニットのC末端とサブユニットのN末端は,成熟型分子の上部に近接していることから,前駆体分子の上部を覆い隠すように独立したドメインとして存在しているペプチドが切り出されて露出する分子の上部表面が活性の発現に重要であると考察している。 さらに,ポリエチレングリコールを結晶化剤として,AS析出結晶とほぼ同型なPEG析出結晶を得て,分解能1.8ÅでのX線結晶構造解析を行っている。これら結晶中のSMK因子の三次元構造は,原子座標の根二乗平均偏差の値が 原子間で0.16Å,全非水素原子間で0.32Åと小さく,ほぼ同一である。しかし,AS析出結晶では塩基性アミノ酸残基の近傍の6ヶ所に硫酸イオンが結合しているのに対して,PEG析出結晶ではこれら硫酸イオンは存在しない。塩基性残基はSMK因子の分子下部に多く局在しており,この硫酸イオンの結合部位が集中している分子底部が同因子の細胞膜に対する作用部位であると考察している。 原子間で0.16Å,全非水素原子間で0.32Åと小さく,ほぼ同一である。しかし,AS析出結晶では塩基性アミノ酸残基の近傍の6ヶ所に硫酸イオンが結合しているのに対して,PEG析出結晶ではこれら硫酸イオンは存在しない。塩基性残基はSMK因子の分子下部に多く局在しており,この硫酸イオンの結合部位が集中している分子底部が同因子の細胞膜に対する作用部位であると考察している。 SMK因子のpH依存的な解離と変性 SMK因子の分子下部に酸性残基も局在し,いくつかはカルボキシル基同士の水素結合を形成している。水素結合を形成して静電的に中性なカルボキシル基がpHの上昇に伴って解離し,静電的に反発してサブユニットの解離をもたらすと考えられる。 解離機構に関するさらに詳細な知見を得るために,エレクトロスプレーイオン化質量分析法(ESI-MS)を用いてSMK因子のpH滴定実験を行った結果,酸性側ではもっぱらヘテロ二量体が検出されるが,pH4.6以上では急激に各サブユニットへの解離が起こることを見出した。ヘテロニ量体のESI-MS多価イオンの分布状態等から,SMK因子は解離の直前まで高次構造を保持すると考えられ,円偏光二色性(CD)スペクトルによっても同様の結果を得た。これらの結果から,SMK因子の解離と変性の過程は,特定のカルボキシル基間の水素結合の切断を端緒として協同的に急激に起こる二状態的な過程であるとしている。また,CDスペクトル測定では,NaCl濃度の変化に伴う二次構造の変化が認められず,塩濃度の上昇は,SMK因子の三次元構造ではなく,その標的細胞の感受性に影響を及ぼすとの可能性を指摘している。 本論文は,耐塩性酵母のSMKキラー因子について,三次元構造をX線解析により明らかにし,その成熟過程,細胞膜との相互作用部位,類縁蛋白質との進化的および機能的な関連性,解離と変性のpH依存,活性の塩濃度依存などについて有用な知見を与えている。よって,本論文は,蛋白質の構造化学,構造生物学および薬学に寄与するところ大であり,博士(薬学)の学位の授与に値する内容を有すると判定した。 |