本研究は、血液細胞における刺激伝達系の研究の多くがマウスやヒトの細胞株で進められ、正常血液細胞における刺激伝達系の知見は少なく、しかも正常ヒト血液細胞での報告は好中球における知見が主で、単球をはじめとする他の細胞での報告は更に少ないことから、正常ヒト血液細胞として単球をとり上げ、サイトカインおよび炎症性メディエーターにより刺激された正常ヒト単球における刺激伝達経路について調べたものであり、下記の結果を得ている。 尚、正常ヒト単球としては、正常ヒト血液より分離した単核球浮遊液よりエリトリエーターを用いて浮遊状態のまま分離して得た単球を用いている。 1.顆粒球マクロファージコロニ-刺激因子(granulocyte-macrophage colony stimulation factor;GM-CSF)及びインターロイキン-3(interleukin-3;IL-3)で正常ヒト単球を刺激すると92kDaの蛋白がチロシンリン酸化されることが認められた。免疫沈降法によりこの92kDaの蛋白は転写因子Stat5であり、ほぼ同様の分子量を持つStat familyの他のメンバーのStat1、Stat3、Stat6ではないこと、更に癌原遺伝子産物のc-Fes(92kDa)やVav(95kDa)でもないことが確認された。実際、この92kDaの蛋白がGM-CSFの刺激により細胞質から核内に移行することをウエスタンブロティング法で確認し、更に、核内移行した92kDaの蛋白がDNAと結合することを、Stat5の認識領域である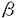 -casein promoter領域の一部(-CAP)をプローブにして、ゲルシフトアッセイを行って確認した。-casein promoter領域に結合した92kDaの蛋白が転写活性を有することは、-CAPの下流にルシフェラーゼ遺伝子を構築したルシフェラーゼレポータープラスミドを正常ヒト単球に導入して、GM-CSF刺激によりルシフェラーゼ活性が有意に上昇することで確認した。 -casein promoter領域の一部(-CAP)をプローブにして、ゲルシフトアッセイを行って確認した。-casein promoter領域に結合した92kDaの蛋白が転写活性を有することは、-CAPの下流にルシフェラーゼ遺伝子を構築したルシフェラーゼレポータープラスミドを正常ヒト単球に導入して、GM-CSF刺激によりルシフェラーゼ活性が有意に上昇することで確認した。 2.菌体成分のn-formyl-methionyl-leucyl-phenylalanine(FMLP)やホルボールエステルのphorbol myristate acetate(PMA)で正常ヒト単球を刺激すると42及び44kDaの蛋白がチロシンリン酸化されることが認められた。免疫沈降法によりこれらの蛋白はmitogen activated protein kinase(MAPK)familyのメンバーであるextracellular signal‐regulated kinase(ERK)であることが確認され、ERKの上流に位置するMAPK/ERK kinase(MEK1)の特異的阻害剤であるPD98059によりFMLPおよびPMAの刺激によるチロシンリン酸化が阻害されることが確かめられた。正常ヒト好中球や、ヒトのGM-CSF依存性である巨核球系細胞株M07eでは、GM-CSFの刺激でもERKがチロシンリン酸化されて活性化されるが、正常ヒト単球ではGM-CSFの刺激によりERKのチロシンリン酸化が誘導されないことを抗ホスホチロシン抗体で確認し、さらに、酵素活性も誘導されないことをERKの特異的基質であるElk-1がリン酸化されないことで確認した。レセプターからERKに至る刺激伝達経路にはShc、Grb2、Sos1、c-Raf-1等の蛋白の存在が知られている。M07eではGM-CSFの刺激によりShcがリン酸化され、Grb2と結合するが、正常ヒト単球ではGM-CSFの刺激でShcがリン酸化されないにもかかわらず、ShcはGrb2と結合した状態で存在すること、M07eではGM-CSFやIL-3及びPMAの刺激によってSos1やc-Raf-1がセリンスレオニンリン酸化されるが、正常ヒト単球ではPMA以外のGM-CSFやIL-3及びFFMLPの刺激ではSos1やc-Raf-1のセリンスレオニンリン酸化が誘導されないことが確かめられた。PMAによる刺激はプロテインキナーゼC(PKC)を介して伝達されてERKが活性化されるが、FMLPによるERKの刺激伝達経路にはPKCは関与せずホスファチジルイノシトール-3-キナーゼ(PI3K)が関与していることが、それぞれPKC阻害剤bisindolylmaleimideやUCN-01、及びPI3K阻害剤wortmanninやLY294002を用いて確かめられた。 3.ERK以外のMAPK familyのメンバーであるp38は、正常ヒト好中球では、GM-CSFの刺激で活性化されないが、tumor necrosis factor(TNF)やFMLPの刺激で強く、PMAの刺激でも弱いながら活性化されることが知られている。これに対し、正常ヒト単球ではGM-CSF、IL-3、TNF、FMLP及びPMAの刺激の有無によらず、常にp38がリン酸化され、活性化されていることを、免疫沈降法、ならびにリン酸化型p38に対する特異的抗体を用いたウエスタンブロティング法、及びp38の特異的基質であるATF-2を用いたin vitroキナーゼアッセイ法で確認した。 4.ERKやp38の生物学的機能における役割を推測するため、正常ヒト単球がFMLPやPMAの刺激によって活性酸素を産生することに着目し、PD98059やp38の特異的阻害剤SB203580が活性酸素産生に影響を及ぼすか調べたところ、FMLPの刺激による活性酸素産生はPD98059により60%に、SB203580により34%にまで減少し、両者同時に用いると13%にまで抑制されることが確認された。これに対し、PMAの刺激による活性酸素産生はPD98059単独では影響を受けず、SB203580による抑制もわずかであったことから、レセプターを介する刺激伝達経路においてERKやp38は重要な位置を占めており、活性酸素産生経路もERKやp38を介していることが示唆された。一方、M07eのGM-CSF依存性の増殖に及ぼす影響をみると、増殖はPD98059により濃度依存性に抑制されることが確認されたことから、細胞増殖においてERKが何らかの役割を担うことが示唆された。 5.正常ヒト単球における刺激伝達経路の主要蛋白含有量を、正常ヒト好中球、U937、HL60、M07eと比較検討した。Jak-Stat系のJak2含有量は、細胞株に比べ非常に少なかったが、これに対してMAPKに至る経路のShc、Grb2、Sos1、c-Raf-1及びMAPK familyのERKやp38、c-Jun N-terminal kinase(JNK)の含有量には極端な差がみられなかった。刺激伝達経路の主要蛋白含有量の差がシグナルの強さの程度差と必ずしも相関しないことより、正常細胞では様々な未知の調節機構が働いていることが示唆された。 以上、本論文はこれまであまり知られていなかった正常ヒト単球における、サイトカインおよび炎症性メディエーターの刺激伝達経路を解明した。正常細胞における刺激伝達経路はいわば基本と言うべき経路であり、この経路の解明により、正常細胞における刺激伝達系との差異が細胞の癌化につながるのではないか等、細胞株での刺激伝達系の研究が更なる意味を持ってくるものと考えられ、このような観点から、学位の授与に値するものと考えられる。 |