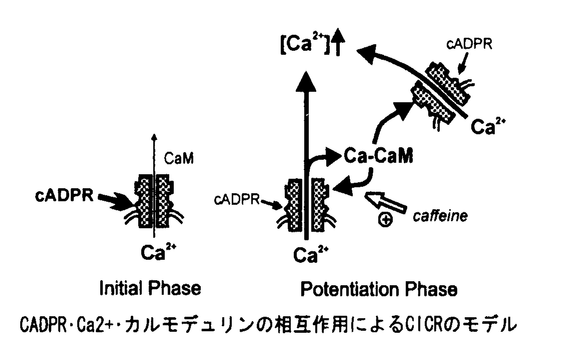
本研究は、ホルモン作用をはじめとする各種の細胞内情報伝達において重要な役割を果たしているものと考えられる自己再生的細胞内カルシウムシグナル発生の一機構を明らかにする目的で、ホルモン応答性細胞とウニ卵の細胞内カルシウムプールに直接コンタクトできる系で、細胞内カルシウムプールの機能的分布と、最近発見された新しい細胞内カルシウム動員物質である環状ADPリボース(cADPR)の作用機構の解析を試みたもので、下記の結果を得ている。 1.まずホルモン応答細胞であるGH4C1細胞においてカルシウム(Ca+)プールの機能的解剖とCa2+平衡状態の解析を試み、GH4C1細胞には 1)Tg感受性Ca2+-ATPaseとInsp3感受性Ca2+チャネルを有する。 2)Tg感受性Ca2+-ATPaseを持つがInsP3感受性Ca2+チャネルを持たない。 3)Tg非感受性Ca2+-ATPaseと、Ca2+誘発性Ca2+放出(CICR)に関連するCa2+チャネルを持つ。 の3つの異なるCa2+プールが存在することを明らかにし、この細胞に自己再生的カルシウムシグナル発生の鍵となりうるCICRプールが存在する可能性を示している。 2.次にウニ卵を用い、CICRプールからのcADPR誘発性Ca2+放出機構を解析している。ここで、ウニ卵におけるcADPRによるCa2+放出反応はCa2+やカフェインの共存により増強される事、などからCa2+そのもの及び何らかの細胞質性増強因子の介在を推測し、この因子がカルモデュリンであることを突き止めている。更に詳細な解析結果をもとにcADPR/Ca2+・カルモデュリン/カフェイン等の相互作用のモデル仮説を立てている。この仮説は、cADPR及びカルモデュリンが予め存在する状態では、添加Ca2+に同期したCa2+放出、即ち真のCICRが起こせることを予測させるものであり、実際Ca2+添加のみでCa2+放出を再現性良く誘発できる事を実験的に示している。 3.この特性からしてcADPRはカルシウム動員物質(セカンドメッセンジャー)と呼ぶよりモデュレーターでありCa2+こそがトリガー兼セカンドメッセンジャーと捉えた方が適切であることを指摘している。この観察・モデルをもとに、今後、このcADPRの作用が更に詳細に解析され、哺乳類細胞での細胞内Ca2+情報伝達、特に自己再生的Ca2+シグナル機構がさらに解明されてゆくものと考えられる。 以上、本論文は、哺乳類非筋細胞の自己再生的Ca2+シグナルにおけるCICRプールの関与の可能性、および、cADPR誘発性CICRの機構の一端を明らかにした。本研究は、これまで詳細が知られていなかった哺乳類の非筋・体細胞における自己再生的カルシウムシグナル発生機構の解明とcADPRのそのシグナルの中での生理的意義の解明に重要な貢献をなすと考えられ、学位の授与に値するものと考えられる。 |