Gas6(product of growth arrest-specific gene 6)は線維芽細胞において細胞増殖停止時に発現が上昇する遺伝子の産物であり、N末端からGla( -カルボキシグルタミン酸)ドメイン、4つのEGF様構造およびC末端ドメインという特徴的な構造を有している。Gas6はC末端ドメインを介してその特異的なレセプターであるAxlと結合し、これまでに知られているGas6の生理活性である細胞増殖促進作用やアポトーシス抑制作用はAxlとの結合およびリン酸化を介している。またレセプターのリン酸化やこれらの生理活性はGlaドメインのないGas6やC末端ドメインのみでも誘導されることも明らかとなった。血液凝固因子などのGlaドメインを有する蛋白質では、Glaドメインはホスファチジルセリン(PS)と結合するのに重要であり、このPSとの結合がそれら蛋白質の活性に不可欠であることが知られている。Gas6はGlaドメインを有しているが、Gas6の細胞増殖促進活性およびアポトーシス抑制活性ではPSへの結合は必要なく、Glaドメインを介してPSに結合することにより発揮されるGas6の新たな機能が存在することが予想された。 -カルボキシグルタミン酸)ドメイン、4つのEGF様構造およびC末端ドメインという特徴的な構造を有している。Gas6はC末端ドメインを介してその特異的なレセプターであるAxlと結合し、これまでに知られているGas6の生理活性である細胞増殖促進作用やアポトーシス抑制作用はAxlとの結合およびリン酸化を介している。またレセプターのリン酸化やこれらの生理活性はGlaドメインのないGas6やC末端ドメインのみでも誘導されることも明らかとなった。血液凝固因子などのGlaドメインを有する蛋白質では、Glaドメインはホスファチジルセリン(PS)と結合するのに重要であり、このPSとの結合がそれら蛋白質の活性に不可欠であることが知られている。Gas6はGlaドメインを有しているが、Gas6の細胞増殖促進活性およびアポトーシス抑制活性ではPSへの結合は必要なく、Glaドメインを介してPSに結合することにより発揮されるGas6の新たな機能が存在することが予想された。 そこでまず、Gas6とリン脂質との相互作用をELISA法を用いて検討したところ、Gas6はPSと結合するが、ホスファチジルコリン(PC)、ホスファチジルエタノールアミン(PE)、ホスファチジルイノシトール(PI)とは結合しなかった。また、この結合にはCa2+およびGla残基が重要であることがわかった。 次に、細胞上のAxlとPSがGas6を介して結合するかをどうかを、Axlを発現しているU937細胞の、リン脂質をコートしたプレートに対する結合により検討した。U937細胞はPC、PEをコートしたプレートには結合しなかったが、PSをコートしたプレートにはCa2+およびGas6依存的に結合した。これらの結果からGas6が分子内の異なる部位でPSおよびAxlと結合し、Axlを有する細胞とPSとを架橋していることが示唆された。 次に、Axlを発現している細胞としてマクロファージを用い、Gas6がマクロファージとPSリポソームを架橋することにより、マクロファージの重要な機能である貪食作用に影響を与えるかどうかを検討した。チオグリコレイトで誘導したマウスの腹腔マクロファージを用いて、蛍光標識PSリポソームの取り込みを蛍光顕微鏡で検出したところ、Gas6非存在下ではわずかしか蛍光が認められないが、Gas6の存在下ではマクロファージがリポソームを細胞内のオルガネラに蓄積しているのが観察された。また、その取り込みはGas6の濃度およびリポソームの濃度に依存したものであった。経時変化を調べたところ、Gas6に依存したPSリポソームの取り込みは1時間以内に活発に起こり、1時間以降ではほとんど起こらないことがわかった(図1)。また、リン脂質特異性を検討した結果、Gas6はPSリポソームの取り込みのみを上昇した(図2)。これまでに報告されているマクロファージによるリポソームの取り込みはゆっくりと、時間に依存して起こり、PS以外にもPIリポソームも取り込むといわれている。これらの結果から、Gas6を介する取り込みは非常に速く起こり、PS特異的であることがわかり、全く性質の違う取り込みであることが明らかとなった。 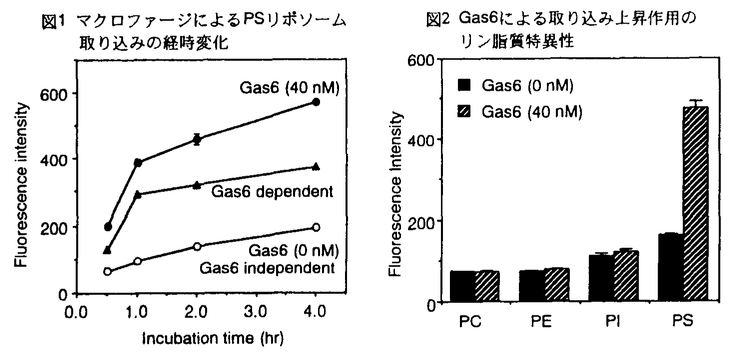 図表図1 マクロファージによるPSリポソーム取り込みの経時変化 / 図2 Gas6による取り込み上昇作用のリン脂質特異性 図表図1 マクロファージによるPSリポソーム取り込みの経時変化 / 図2 Gas6による取り込み上昇作用のリン脂質特異性 次にGas6がPSリポソームの取り込みを上昇させるメカニズムを検討した。まず、Gas6の取り込み上昇作用におけるGas6とマクロファージの結合の重要性を検討した。Gas6のレセプター結合部位に結合し、Gas6とレセプターとの結合を抑制すると考えられるAxlの細胞外ドメインを加えると、Gas6によるPSリポソームの取り込み上昇作用がほぼ完全に阻害されることが示され,Gas6のレセプター結合部位とマクロファージとの結合がPSリポソームの取り込み上昇に不可欠であることが示唆された(図3)。次に,Gas6とPSリポソームの結合の重要性を検討した。Axlへの結合性、リン酸化能は保持しており、PSとの結合性が失われていると考えられるGlaドメインのないGas6[(-Gla Domain)Gas6]を調製した。予想どおりGlaドメイン欠失Gas6はPSとの結合性を失っていたが、Axlのリン酸化能は有していた。このGlaドメイン欠失Gas6の存在下でマクロファージによるPSリポソームの取り込みを検討したところ、取り込みが全く上昇しないことがわかった。このことからGas6のPSリポソーム取り込み上昇作用にはGas6とPSリポソームとの結合が重要であり、Axlのリン酸化により別のレセプターによる取り込みが上昇しているのではないことが明らかになった。  図3 Gas6のPSリポソーム取り込み上昇作用のAxl細胞外ドメインによる阻害 図3 Gas6のPSリポソーム取り込み上昇作用のAxl細胞外ドメインによる阻害 胸腺などのアポトーシスが頻繁に起こっている組織中にマクロファージはわずかしか存在しておらず、マクロファージ以外の細胞がアポトーシス細胞を貪食し、除去している可能性が考えられる。例えば血管平滑筋細胞や血管内皮細胞がアポトーシスした同種細胞を取り込むことが報告されている。ラット血管平滑筋細胞におけるPSリポソームの取り込みを検討したところ、Gas6により約1.5倍に上昇し、Axlの細胞外ドメインによりこの上昇は完全に抑制された。このことからGas6はマクロファージ以外の細胞においてもマクロファージと同様の機構により貪食を促進していることが示された。 PSは正常では生体膜の脂質二重層の内側にほとんどすべてが局在しているが、アポトーシス細胞や老化赤血球、活性化した血小板では表面に存在することが報告されている。それゆえに、PSは除去されるべき細胞をマクロファージが認識する際のマーカーの一つであると考えられている。そこで、Gas6がマクロファージによるアポトーシス細胞の認識、取り込みを上昇させるかを検討した。まず、アポトーシスを誘導したマウス胸腺細胞へのGas6の結合をフローサイトメトリーを用いて検討した。アポトーシスを誘導するとアポトーシス細胞のマーカーとして使われているアネキシンVで染色される細胞数が増加していることがわかった。これはアポトーシスがこの条件で効率よく誘導されていることを示している。Gas6と結合性を示す細胞もアポトーシスの誘導とともに増加した。このことからGas6がアポトーシス細胞に結合することが示された。 次に、Gas6がマクロファージによるアポトーシス細胞の取り込みに影響を与えるかどうかを検討した。51Crでラベルしたマウス胸腺細胞にアポトーシスを誘導し、マクロファージによる取り込みを検出した。その結果、Gas6存在下ではマクロファージによるアポトーシス細胞の取り込みが約2倍に上昇し(図4)。このことから、Gas6がマクロファージによるアポトーシス細胞の取り込みも上昇させる作用を有していることが明らかとなった。  図4 Gas6のアポトーシス細胞取り込み増強作用 図4 Gas6のアポトーシス細胞取り込み増強作用 以上の結果により、Gas6存在下においてマクロファージによるPSリポソームの取り込みが上昇し、この上昇にはGas6とマクロファージとの結合およびGas6とPSリポソームの結合がともに必要であった。これはGas6がGlaドメインを介してPSリポソームと、C末端ドメインを介してマクロファージと結合し、PSリポソームとマクロファージを架橋することによるものと考えられる。これまでに報告されているマクロファージによるリポソームの取り込みと比較すると、1時間という早い時間で取り込まれ、PSリポソーム特異的であるという特徴があり、性質の異なるものであった。また、Gas6はマクロファージによるアポトーシス細胞の取り込みも上昇させた。今回の結果からはGas6がマクロファージ上のどの分子と結合しているかは明らかではないが、マクロファージがGas6のレセプターであるAxlを発現していること、Gas6はレセプター結合部位を介してマクロファージと特異的に結合しているという結果から、Axlであると考えられる。 発生や分化、生体の恒常性の維持などの場面で、貪食細胞が不要となった細胞や有害な細胞を取り込んで除去すると考えられている。貪食細胞は除去されるべき細胞の表面上の変化を特異的なレセプターを介して認識するといわれており、特にPSを認識するいくつかのレセプターが関与していることが示された。しかし、すべての貪食をこれらのレセプターで説明することはできず、レセプター以外の機構も関与する可能性が考えられた。今回の研究により、可溶性の蛋白質であるGas6が貪食の一部の役割を担っている可能性が示唆された。生体防御において、PSを露出した細胞をより迅速に除去することは重要であることから、Gas6が血管や精巣、胸腺などの存在部位において不要となった細胞を素早く除去することにより、生体の恒常性の維持の一翼を担っている可能性が考える。今後はGas6の生体内での役割について検討を進めていきたいと考えている。 |